




Abstract
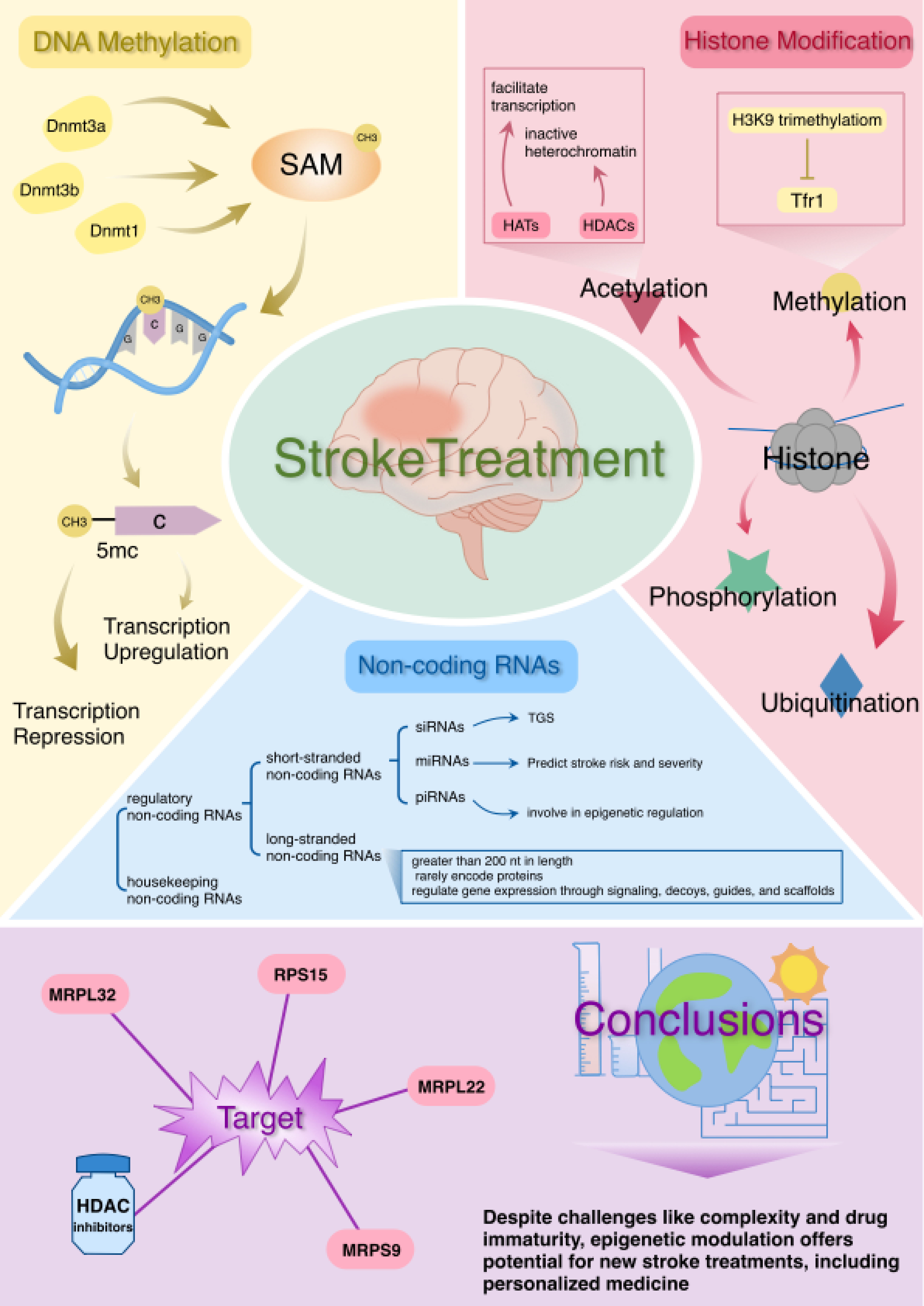
Cerebrovascular diseases are a serious threat to human health, among which stroke is extremely harmful. In recent years, more and more studies have shown that epigenetic regulation is crucial in the development of cerebrovascular diseases. In this review, we focus on the research progress of epigenetic regulation in stroke treatment, detailing the regulatory mechanisms of DNA methylation, histone modification and non-coding RNAs, and analyzing their roles in the pathophysiology of stroke. It was found that ischemic stroke causes dynamic changes in DNA methylation, which affects gene expression and alters the process of injury and recovery; histone modification levels are also altered after stroke, which affects chromatin status and gene transcription; and noncoding RNAs, such as miRNAs, siRNAs, piRNAs, and lncRNAs, which play a key role in gene expression regulation, are associated with the risk of stroke, severity and symptom onset. In addition, therapeutic strategies targeting epigenetic regulation are also discussed. Although facing challenges such as complex mechanisms, susceptibility to environmental influences, and the early stage of drug development, epigenetic regulation is still very promising in the treatment of stroke, and it is expected to provide a new theoretical basis and research direction for the prevention and treatment of stroke in the future.
Keywords:Epigenetic regulation; Stroke; DNA methylation; Histone modification; Non-coding RNA; Therapeutic targets
Introduction
In 2021, stroke maintained its status as the second most prevalent cause of death globally, surpassed only by ischaemic heart disease, and occupied the third spot in terms of Disability-Adjusted Life Years lost due to noncommunicable diseases , trailing behind ischaemic heart disease and neonatal disorders[11] .
Cerebrovascular diseases are lesions of blood vessels in the brain, including cerebral infarction, cerebral hemorrhage, and subarachnoid hemorrhage, which will lead to stroke[72] . In recent years, epigenetic regulation has played an important role in stroke pathogenesis and stroke repair as a higher-order regulatory mechanism for tissue repair. Epigenetic regulation refers to mechanisms that regulate gene expression through non-DNA sequence changes (DNA methylation, histone modifications, and non-coding RNAs)[44] .These changes can affect multiple aspects of gene transcription, splicing, RNA stability and translation without altering the DNA sequence[29] . Epigenetic regulation is a key post-transcriptional regulation of gene expression and plays an important regulatory role in organogenesis, homeostasis and pathology[67] .
In cerebrovascular diseases, alterations in epigenetic regulation may lead to neuronal damage and abnormal responses of vascular endothelial cells. Therefore, investigating the role of epigenetic regulation in cerebrovascular diseases can help us better understand the pathogenesis of these diseases and find new therapeutic strategies.
Epigenetic regulation
DNA methylation
DNA methylation is the process by which methyl groups are added to the DNA molecule. This modification can occur at different locations in the genome and plays an important role in gene regulation. Several enzymes are involved in DNA methylation, including DNA methyltransferases (dnmt) such as Dnmt3a, Dnmt3b, and Dnmt1. DNMTs catalyze the transfer of methyl groups from S-adenosylmethionine (SAM) to cytosine residues in DNA to form 5-methylcytosine (5mC). However, post-mitotic neurons in the mature mammalian brain still express a large number of Dnmts, raising the possibility that Dnmts and DNA methylation may play new roles in the brain[18,24] .
DNA methylation can influence the pathogenesis of stroke by modulating genetic or environmental risk factors. DNA methylation regulation is shaped by two opposing processes, the addition and removal of the fifth methyl group of cytosine in DNA[31,33] . It causes transcriptional repression or transcriptional upregulation by preventing RNA polymerase from recognizing specific DNA regions or by eliminating “DNA methylation breaks”, respectively. The effect of DNA methylation on the expression of transcripts depends on their location in the gene region (promoter and or gene body), and transcriptional changes are usually dependent on methylation at multiple locations in a given gene[36,37] .DNA methylation can be a biomarker for stroke[40] .
Zhan, L. et al. demonstrated that the Piwil2/piRNAs pathway plays a role in the epigenetic regulation of CREB2 expression and neuroprotection of tGCI by HPC in rats. Specifically, HPC-induced downregulation of Piwil2 in rat CA1 inhibited CREB2 hypermethylation and promoted CREB2 transcription by retraining DNMT3A to induce promoter site CpG methylation, leading to an increase in CREB2 after tGCI[75] .
Histone modifications
Histone modifications include several types, each with a specific function. Histone methylation can occur on the Lys and Arg residues of histones and has the effect of reducing the positive charge carried by histones and directly or indirectly affecting the transcriptional activity of genes. Histone methylation is usually associated with gene activation or gene silencing[5] . Histone acetylation occurs predominantly and diversely on core histones. The degree of acetylation is higher in the structural domains of DNA containing active genes. Acetylation/deacetylation modifications can regulate the level of gene transcription, participate in DNA repair, splicing and replication, chromosome assembly, and cellular signaling[61] .Histone phosphorylation is involved in gene transcription, DNA repair, apoptosis and chromosome condensation[27] . Histone ubiquitination is involved in X chromosome inactivation and affects histone methylation and gene transcription[70] .
Histone acetyltransferases (HATs) acetylate lysine residues in histone tails. This promotes DNA unfolding and chromatin deconcentration, which facilitates transcription and protein synthesis. Histone deacetylases remove acetyl groups from histones. This leads to the formation of transcriptionally inactive heterochromatin in which gene expression is blocked. To maintain the transcriptionally active state of chromatin, HATs and hdac need to work in concert[30,58] .
H3K9 trimethylation is a specific methylation modification of the 9th amino acid lysine of histone 3 that inhibits the key transferrin receptor (Tfr1), which determines whether or not a neuron will experience iron death. By decreasing H3K9 trimethylation protects neurons from iron death, whereas increasing it leads to neuronal iron death[42] .
non-coding RNA
Non-coding RNAs that are not translated into proteins can be categorized into housekeeping non-coding RNAs and regulatory non-coding RNAs. RNAs with regulatory roles are divided into two main categories according to their size: short-stranded non-coding RNAs (including siRNAs, miRNAs and piRNAs) and long-stranded non-coding RNAs (LncRNAs)[63,74] .In recent years, a large number of studies have shown that non-coding RNAs play an important role in epigenetic modifications, which can regulate expression at the gene level and chromosome level and control cell differentiation[2,12] .
siRNA is derived from long double-stranded RNA molecules (including RNA produced by viral replication, transposon activity, or gene transcription) that can be cleaved by Dicer's enzyme into 19-24 nt (nt: nucleotide) RNA fragments, which are then loaded onto the Argonaute (AGO) protein to perform their function[34,52] .
miRNAs are single-stranded RNAs of approximately 19-24 nt, 50% of which are located in chromosomal regions prone to structural changes[57] .Initially, it was thought that there were two main differences between siRNAs and miRNAs as a class of regulatory RNAs. One is that miRNAs are endogenous, being the products of biological gene expression, whereas siRNAs are exogenous, originating from viral infections, gene transfer points, or gene targets. The other difference is that miRNAs consist of incomplete hairpin-shaped double-stranded RNAs processed by Drosha and Dicer, whereas siRNAs are products of fully complementary long double-stranded RNAs processed by Dicer[9] .Despite these differences, it has been hypothesized that miRNAs and siRNAs have similar mechanisms of action in mediating transcriptional gene silencing because miRNAs and siRNAs are closely related, e.g., the two fragments are similar in size. Recently, nearly 1,800 putative miRNAs have been identified in the human genome, and the number of miRNAs continues to increase rapidly due to the development of high-throughput sequencing technologies[26,41] .
piRNAs are a class of RNA molecules approximately 26-31 nt in length. Its name piRNA (Piwi-interacting RNA) reflects the fact that piRNA binds to Piwi proteins under physiological conditions.The role of Piwi proteins as epigenetic regulators is reflected by their silencing of the homeobox gene by binding to PcGs (polycistronic ribosomal histones) to genomic PcG response elements. Thus, it has been hypothesized that piRNAs associated with Piwi proteins should also play an important role in epigenetic regulation[45] .
LncRNAs are another class of non-coding regulatory RNAs. lncRNAs are generally greater than 200 nt in length, are located in the nucleus or cytoplasm, and rarely encode proteins.[8,51] .LncRNAs are another class of non-coding regulatory RNAs. lncRNAs are generally greater than 200 nt in length, are located in the nucleus or cytoplasm, and rarely encode proteins[55] .However, these five major categories mainly involve only four typical mechanisms by which lncRNAs regulate gene expression: signaling, decoys, guides, and scaffolds[65] .
Non-coding rna (ncRNAs) are key regulators of gene expression, and the most studied ncRNAs are long-stranded non-coding rna (lncRNAs), micro rna (miRNAs), and circular rna (circRNAs)[66] .For ischemic stroke, the search for biomarkers is crucial for early diagnosis and guiding treatment. Changes in ncRNA expression profiles during ischemic stroke have gradually attracted attention[10,22] . The relative stability, specificity, and reproducibility of ncRNAs make them promising biomarkers for early disease recognition[46] .
Table 1. Comparison of Epigenetic Regulation Types and Mechanisms
| Regulation Type | Specific Modification | Mechanism | Impact on Gene Expression |
|
DNA Methylation |
Methyl group addition |
By DNMTs |
Promoter: often represses; Gene body: varies |
|
Histone Modification |
Methylation |
Alters chromatin |
Activation or silencing |
|
|
Acetylation |
HATs add, HDACs remove |
HATs promote, HDACs inhibit |
|
Non - coding RNA |
siRNA |
With AGO, TGS |
Silences genes |
|
|
miRNA |
Bind mRNA |
Negatively regulates |
|
|
lncRNA |
4 mechanisms |
Multi - level regulation |
Epigenetic regulation and stroke
Association of Epigenetic Modulation with Stroke
Ischemic Stroke Leads to Dynamic Changes in DNA Methylation and Regulates Widespread Differential Gene Expression, Altering Injury and Recovery Processes[3,13] .
Whole blood whole genome methylation levels were higher in adults with ischemic smog disease than in healthy individuals. A total of 759 methylation probes were significantly different between cases and controls. The hypermethylated regions were mainly concentrated in gene spacer regions. Among the genes with the highest degree of differential expression, KCNMA1 and GALNT2 were upregulated and SOX6 and RBM33 were downregulated[28] .
Hypomethylation of TNF Receptor-Related Factor 3 (TRAF3), Hypermethylation of thrombospondin-1 (THBS1), and Increased DNMT3A Activity Also Predict Stroke Outcome and the Extent of Ischemic Injury[13,20] .To date, it has been reported that within the BBB, hypermethylation of the TIMP2 promoter or hypomethylation of the VCAM-1 promoter controls BBB permeability and leukocyte recruitment[54,60] .
Ischemic stroke typically reduces overall gene expression levels by inhibiting acetylation of histones H3 and H4[32,43] . Ischemic stroke overall reduced histone acetylation levels in the ischemic core, penumbral regions, and hemispheres of the affected brain. Acetylation of histone H3 lysine 9 (H3K9Ac) in the ischemic penumbral region of rat cerebral cortex decreased 2-fold at 1 hour and more than 4-fold at 4 and 24 hours after photothrombotic stroke[16] . In addition, Demyanenko's study showed that acetylation levels of histone H4 in the ipsilateral cortical penumbra were either increased or unaffected 7 days after ischemic stroke. Interestingly, the increased acetylation levels returned to control levels 14 days after ischemic stroke[14,15] . Histone H3 and H4 acetylation levels were reduced in oligodendrocytes of cerebral white matter 7 days after ischemic stroke. The acetylation level of histone H4 in the ipsilateral cortex was not altered at 7 or 14 days after ischemic stroke[68] .
It has been shown that ICH significantly reduced the acetylation level of histone H4 in the ipsilateral cortex, and that sodium butyrate (NaB), an HDAC inhibitor, increased the acetylation level and eliminated the decrease in acetylation level caused by ICH. In addition, HDAC inhibitors slightly ameliorated dyskinesia after ICH, and exercise increased the acetylation levels of histones (H3 and H4) in bilateral cortex; interestingly, no synergistic effect of exercise and NaB administration on histone acetylation was observed[48] .
RNA modifications are an important part of epigenetic modifications. Currently, more than 160 chemical modifications have been identified in RNA, which are involved in regulating the structural properties of RNA or altering the affinity of mRNA for ribosomes[76] .miRNAs can predict the risk and severity of stroke and the onset of cerebral infarction symptoms, and up- or down-regulation of multiple miRNAs, rather than a single miRNA, occurs during stroke[1,62] .
Table 2. Epigenetic Changes Related to Stroke and Their Associations
| Stroke Type | Epigenetic Changes | Key Genes/Molecules | Main Impact |
|
Ischemic Stroke |
DNA meth. change; histone acetylation↓; ncRNA expression change |
KCNMA1, SOX6; H3K9Ac; miRNAs |
Affects gene expression, recovery, BBB |
|
ICH |
Histone H4 acetylation↓ in ipsilateral cortex |
Histone H4 |
Affects neuronal survival |
Clinical studies and experimental results
Recent studies have shown that siRNAs can lead to cellular transcriptional gene silencing (TGS) through DNA methylation and histone modification[7,39] . Zhou et al. demonstrated that siRNA silencing of EZH2 reversed cisplatin resistance in human non-small cell lung cancer and gastric cancer cells[77] . EZH2 acts as a histone methyltransferase that causes methylation of H3K27, which then serves as an anchor for CpG methylation, leading to the formation of silent chromatin and ultimately TGS[64] .Chromatin immunoprecipitation experiments showed that the binding of EZH2-inhibited DNMTs to specific genes depends on the presence of EZH2. Bisulfite sequencing results also demonstrated that EZH2 is required for methylation of target promoters repressed by EZH2, suggesting that EZH2 is involved in DNA methylation by recruiting DNA methyltransferases[71] .
Dysregulation of histone methylation has been linked to a variety of diseases, including cancer, neurodegenerative diseases and genetic disorders. Histone methylation marks are also thought to be inherited through mitosis and meiosis, which provides a potential mechanism for transgenerational transmission of epigenetic information[6] .
Many histone methyltransferases and demethyltransferases have been identified that catalyze the addition and removal of methyl groups from histone tails. These enzymes are key regulators of histone methylation dynamics, which in turn control the transcription of specific genes and gene networks[17] .Histone methyltransferases commonly act as oncogenes in cancer, promoting cell proliferation and survival[50] ,and Histone Demethylases Are Associated with Neurodegenerative Diseases and Age-Related Cognitive Decline[49] .
The exploration of epigenetic mechanisms in Parkinson's disease (PD) has witnessed significant advancements, with a particular focus on the role of DNA methylation in the context of PD. A study on a Sudanese cohort has provided valuable insights into this complex relationship.In this clinical investigation, researchers gathered a cohort of 172 Sudanese individuals, with 90 diagnosed with PD and 82 serving as healthy controls. An interesting aspect was the relatively young age of onset in the PD patients, averaging 40.6 ± 22.4 years, and a notable 64 patients had a family history of the disease.The study's findings regarding the methylation of the SNCA gene were quite revealing. Among the analyzed CpG sites, CpGs 16 - 23 exhibited significant hypomethylation in PD patients when compared to the control group. The P - values, ranging from 0.023 to 0.003, indicated a strong association. When sporadic cases were excluded from the analysis, the significance of these results was further enhanced. This suggests that the hypomethylation of these specific CpG sites might be a crucial epigenetic factor contributing to the development of PD, especially in familial cases[4] .
Aberrant histone methylation has been reported in a variety of cancers, with global loss and gain of specific histone methylation marks observed. These changes are thought to promote dysregulation of gene expression programs for cancer development by driving cellular transformation and metastasis. Histone methylation is also involved in DNA repair, where histone H3K36 methylation is a key signal for mismatch repair. Defects associated with histone methylation and DNA repair have been linked to hereditary diseases such as hereditary nonpolyposis colorectal cancer and xeroderma pigmentosum[35,56] .
Epigenetic regulation as a therapeutic target
Histone methylation is a dynamic epigenetic mark in health, disease, and heredity, and understanding its regulation and function is critical to understanding normal biology and disease etiology[25] . A number of small molecule inhibitors of histone methyltransferases and demethyltransferases have been developed, and these inhibitors are currently being evaluated in preclinical trials for the treatment of cancer and other diseases[50] . Histone acetylation is also a target for epigenetic therapies, and drugs such as histone deacetylase inhibitors show promise in the treatment of neurodegenerative diseases and other disorders caused by aberrant histone acetylation[21] .
Fujii et al. showed that daily consumption of large amounts of vegetables reduced ABCA1 gene methylation and lowered cholesterol and atherosclerosis. But interestingly, only women validated this study[19] .
Exercise and HDAC inhibitors can alter epigenetic modifications in the motor cortex. These changes include increased DNA methylation, decreased histone deacetylase activity, and increased histone acetylation levels. These modifications can affect gene expression and promote neuronal plasticity and anti-inflammatory responses[48] .It has been shown that in an ischemic stroke model, administration of a histone deacetylase inhibitor (HDACi) improves motor and cognitive function after stroke, with concomitant neuroplasticity, neurogenesis, and attenuated inflammation[32,38] .
A researcher conducted a PPI network analysis of differential miRNAs, differential genes, and differentially methylated sites, and the results of the study showed that MRPS9, MRPL22, MRPL32, and RPS15 were identified as potential diagnostic and therapeutic targets for ischemic stroke progression. This implies that these genes may be involved in the process of ischemic stroke onset and progression, and therefore could be potential therapeutic targets[78] .
Hypomethylation and altered expression of the miR-223 gene may lead to its inactivation or reduced degradation at the post-transcriptional level, which increases its inhibitory effect on target genes, and consequently contributes to the development and progression of stroke risk factors such as atherosclerosis[59] .
Table 3. Epigenetic Regulation as Therapeutic Targets for Stroke and Related Drug Development
| Regulation Type | Therapeutic Target | Interventions | Research Stage |
|
DNA Methylation |
Gene methylation sites |
Diet - based |
Clinical obs. |
|
Histone Modification |
Methyl/demethyl - transferases; HDACs |
Small - molecule inhibitors; HDAC inhibitors |
Pre - clinical; some clinical (HDACi) |
|
Non - coding RNA |
Differential miRNA/lncRNA pathways |
Antisense oligonucleotides, mimics |
Pre - clinical |
Conclusion
Epigenetic modulation as a therapeutic strategy for stroke also faces several challenges. First, epigenetic mechanisms are complex and involve the interaction of multiple factors. This makes therapeutic strategies targeting specific epigenetic modifications difficult. Second, epigenetic modifications may be influenced by a variety of factors such as environment and lifestyle, which increases the difficulty of implementing effective interventions in stroke patients. In addition, most of the current epigenetic drugs are in their early stages and require further research and clinical trials to validate their safety and efficacy.
Despite the challenges, epigenetic modulation holds great promise in stroke treatment. First, by studying epigenetic mechanisms, we can gain a deeper understanding of stroke onset and progression. This can help discover new therapeutic targets and predictive biomarkers. Second, epigenetic drugs have the potential to improve stroke symptoms and prognosis by regulating specific gene expression. For example, neuronal damage after stroke can be reduced by upregulating the expression of genes with protective effects. Finally, epigenetic therapies can target individual differences for precision medicine and provide more personalized treatment plans for stroke patients.
Acknowledgements
Not applicable.
Author contributions
All authors were involved in the conceptualization and design of the study. Data collection, first draft of the manuscript, and translation were done by Tiancai Yang. All authors commented on a previous version of the manuscript. All authors read and approved the final manuscript.
Consent for publication
Not applicable.
Funding information
This study was supported by the Jining City Key R&D Program Project (2022YXNS137).
Competing Interests
The authors declare that they have no existing or potential commercial or financial relationships that could create a conflict of interest at the time of conducting this study.
Data Availability
The data that support the findings of this study are available on request from the corresponding author.
References
[1] Abe A, Tanaka M, Yasuoka A, Saito Y, Okada S, Mishina M, et al. (2020) Changes in Whole-Blood microRNA Profiles during the Onset and Treatment Process of Cerebral Infarction: A Human Study. Int J Mol Sci, 21(9). https://doi.org/10.3390/ijms21093107
[2] Amaral PP, Dinger ME, Mercer TR, Mattick JS. (2008) The eukaryotic genome as an RNA machine. Science, 319(5871)1787-9. https://doi.org/10.1126/science.1155472
[3] Baccarelli A, Tarantini L, Wright RO, Bollati V, Litonjua AA, Zanobetti A, et al. (2010) Repetitive element DNA methylation and circulating endothelial and inflammation markers in the VA normative aging study. Epigenetics, 5(3)222-8. https://doi.org/10.4161/epi.5.3.11377
[4] Bakhit Y, Schmitt I, Hamed A, Ibrahim EAA, Mohamed IN, El-Sadig SM, et al. (2022) Methylation of alpha-synuclein in a Sudanese cohort. Parkinsonism Relat Disord, 1016-8. https://doi.org/10.1016/j.parkreldis.2022.05.009
[5] Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, et al. (2007) High-resolution profiling of histone methylations in the human genome. Cell, 129(4)823-37. https://doi.org/10.1016/j.cell.2007.05.009
[6] Basavarajappa BS, Subbanna S. (2021) Histone Methylation Regulation in Neurodegenerative Disorders. Int J Mol Sci, 22(9). https://doi.org/10.3390/ijms22094654
[7] Bayne EH, Allshire RC. (2005) RNA-directed transcriptional gene silencing in mammals. Trends Genet, 21(7)370-3. https://doi.org/10.1016/j.tig.2005.05.007
[8] Cao X, Yeo G, Muotri AR, Kuwabara T, Gage FH. (2006) Noncoding RNAs in the mammalian central nervous system. Annu Rev Neurosci, 2977-103. https://doi.org/10.1146/annurev.neuro.29.051605.112839
[9] Carthew RW, Sontheimer EJ. (2009) Origins and Mechanisms of miRNAs and siRNAs. Cell, 136(4)642-55. https://doi.org/10.1016/j.cell.2009.01.035
[10] Chen J, Liu P, Dong X, Jin J, Xu Y. (2021) The role of lncRNAs in ischemic stroke. Neurochem Int, 147105019. https://doi.org/10.1016/j.neuint.2021.105019
[11] Collaborators GBDSRF. (2024) Global, regional, and national burden of stroke and its risk factors, 1990-2021: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Neurol, 23(10)973-1003. https://doi.org/10.1016/S1474-4422(24)00369-7
[12] Costa FF. (2008) Non-coding RNAs, epigenetics and complexity. Gene, 410(1)9-17. https://doi.org/10.1016/j.gene.2007.12.008
[13] Cullell N, Soriano-Tarraga C, Gallego-Fabrega C, Carcel-Marquez J, Torres-Aguila NP, Muino E, et al. (2022) DNA Methylation and Ischemic Stroke Risk: An Epigenome-Wide Association Study. Thromb Haemost, 122(10)1767-78. https://doi.org/10.1055/s-0042-1749328
[14] Demyanenko S, Berezhnaya E, Neginskaya M, Rodkin S, Dzreyan V, Pitinova M. (2019) capital ES, Cyrilliclass II histone deacetylases in the post-stroke recovery period-expression, cellular, and subcellular localization-promising targets for neuroprotection. J Cell Biochem, 120(12)19590-609. https://doi.org/10.1002/jcb.29266
[15] Demyanenko S, Neginskaya M, Berezhnaya E. (2018) Expression of Class I Histone Deacetylases in Ipsilateral and Contralateral Hemispheres after the Focal Photothrombotic Infarction in the Mouse Brain. Transl Stroke Res, 9(5)471-83. https://doi.org/10.1007/s12975-017-0595-6
[16] Demyanenko S, Uzdensky A. (2019) Epigenetic Alterations Induced by Photothrombotic Stroke in the Rat Cerebral Cortex: Deacetylation of Histone h3, Upregulation of Histone Deacetylases and Histone Acetyltransferases. Int J Mol Sci, 20(12). https://doi.org/10.3390/ijms20122882
[17] Deng P, Chen QM, Hong C, Wang CY. (2015) Histone methyltransferases and demethylases: regulators in balancing osteogenic and adipogenic differentiation of mesenchymal stem cells. Int J Oral Sci, 7(4)197-204. https://doi.org/10.1038/ijos.2015.41
[18] Feng J, Chang H, Li E, Fan G. (2005) Dynamic expression of de novo DNA methyltransferases Dnmt3a and Dnmt3b in the central nervous system. J Neurosci Res, 79(6)734-46. https://doi.org/10.1002/jnr.20404
[19] Fujii R, Yamada H, Munetsuna E, Yamazaki M, Mizuno G, Tsuboi Y, et al. (2019) Dietary vegetable intake is inversely associated with ATP-binding cassette protein A1 (ABCA1) DNA methylation levels among Japanese women. Nutrition, 651-5. https://doi.org/10.1016/j.nut.2019.02.010
[20] Gallego-Fabrega C, Carrera C, Reny JL, Fontana P, Slowik A, Pera J, et al. (2016) TRAF3 Epigenetic Regulation Is Associated With Vascular Recurrence in Patients With Ischemic Stroke. Stroke, 47(5)1180-6. https://doi.org/10.1161/STROKEAHA.115.012237
[21] Gediya P, Parikh PK, Vyas VK, Ghate MD. (2021) Histone deacetylase 2: A potential therapeutic target for cancer and neurodegenerative disorders. Eur J Med Chem, 216113332. https://doi.org/10.1016/j.ejmech.2021.113332
[22] Ghafouri-Fard S, Shoorei H, Taheri M. (2020) Non-coding RNAs participate in the ischemia-reperfusion injury. Biomed Pharmacother, 129110419. https://doi.org/10.1016/j.biopha.2020.110419
[23] Ghildiyal M, Zamore PD. (2009) Small silencing RNAs: an expanding universe. Nat Rev Genet, 10(2)94-108. https://doi.org/10.1038/nrg2504
[24] Goto K, Numata M, Komura JI, Ono T, Bestor TH, Kondo H. (1994) Expression of DNA methyltransferase gene in mature and immature neurons as well as proliferating cells in mice. Differentiation, 56(1-2)39-44. https://doi.org/10.1046/j.1432-0436.1994.56120039.x
[25] Greer EL, Shi Y. (2012) Histone methylation: a dynamic mark in health, disease and inheritance. Nat Rev Genet, 13(5)343-57. https://doi.org/10.1038/nrg3173
[26] Griffiths-Jones S, Grocock RJ, van Dongen S, Bateman A, Enright AJ. (2006) miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res, 34(Database issue)D140-4. https://doi.org/10.1093/nar/gkj112
[27] Gurley LR, Walters RA, Tobey RA. (1973) Histone phosphorylation in late interphase and mitosis. Biochem Biophys Res Commun, 50(3)744-50. https://doi.org/10.1016/0006-291x(73)91307-7
[28] He S, Ye X, Duan R, Zhao Y, Wei Y, Wang Y, et al. (2022) Epigenome-Wide Association Study Reveals Differential Methylation Sites and Association of Gene Expression Regulation with Ischemic Moyamoya Disease in Adults. Oxid Med Cell Longev, 20227192060. https://doi.org/10.1155/2022/7192060
[29] Holliday R. (2006) Epigenetics: a historical overview. Epigenetics, 1(2)76-80. https://doi.org/10.4161/epi.1.2.2762
[30] Hu Z, Zhong B, Tan J, Chen C, Lei Q, Zeng L. (2017) The Emerging Role of Epigenetics in Cerebral Ischemia. Mol Neurobiol, 54(3)1887-905. https://doi.org/10.1007/s12035-016-9788-3
[31] Ito S, D'Alessio AC, Taranova OV, Hong K, Sowers LC, Zhang Y. (2010) Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature, 466(7310)1129-33. https://doi.org/10.1038/nature09303
[32] Jhelum P, Karisetty BC, Kumar A, Chakravarty S. (2017) Implications of Epigenetic Mechanisms and their Targets in Cerebral Ischemia Models. Curr Neuropharmacol, 15(6)815-30. https://doi.org/10.2174/1570159X14666161213143907
[33] Jiang D, Wang Y, Shen Y, Xu Y, Zhu H, Wang J, et al. (2016) Estrogen and promoter methylation in the regulation of PLA2G7 transcription. Gene, 591(1)262-7. https://doi.org/10.1016/j.gene.2016.07.048
[34] Jinek M, Doudna JA. (2009) A three-dimensional view of the molecular machinery of RNA interference. Nature, 457(7228)405-12. https://doi.org/10.1038/nature07755
[35] Joensuu EI, Nieminen TT, Lotsari JE, Pavicic W, Abdel-Rahman WM, Peltomaki P. (2015) Methyltransferase expression and tumor suppressor gene methylation in sporadic and familial colorectal cancer. Genes Chromosomes Cancer, 54(12)776-87. https://doi.org/10.1002/gcc.22289
[36] Jones PA. (2012) Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet, 13(7)484-92. https://doi.org/10.1038/nrg3230
[37] Jones PA, Baylin SB. (2007) The epigenomics of cancer. Cell, 128(4)683-92. https://doi.org/10.1016/j.cell.2007.01.029
[38] Kassis H, Shehadah A, Chopp M, Zhang ZG. (2017) Epigenetics in Stroke Recovery. Genes (Basel), 8(3). https://doi.org/10.3390/genes8030089
[39] Kawasaki H, Taira K. (2004) Induction of DNA methylation and gene silencing by short interfering RNAs in human cells. Nature, 431(7005)211-7. https://doi.org/10.1038/nature02889
[40] Kim M, Long TI, Arakawa K, Wang R, Yu MC, Laird PW. (2010) DNA methylation as a biomarker for cardiovascular disease risk. PLoS One, 5(3)e9692. https://doi.org/10.1371/journal.pone.0009692
[41] Kozomara A, Griffiths-Jones S. (2014) miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res, 42(Database issue)D68-73. https://doi.org/10.1093/nar/gkt1181
[42] Lan T, Hu L, Sun T, Wang X, Xiao Z, Shen D, et al. (2023) H3K9 trimethylation dictates neuronal ferroptosis through repressing Tfr1. J Cereb Blood Flow Metab, 43(8)1365-81. https://doi.org/10.1177/0271678X231165653
[43] Lanzillotta A, Pignataro G, Branca C, Cuomo O, Sarnico I, Benarese M, et al. (2013) Targeted acetylation of NF-kappaB/RelA and histones by epigenetic drugs reduces post-ischemic brain injury in mice with an extended therapeutic window. Neurobiol Dis, 49177-89. https://doi.org/10.1016/j.nbd.2012.08.018
[44] Li Y. (2021) Modern epigenetics methods in biological research. Methods, 187104-13. https://doi.org/10.1016/j.ymeth.2020.06.022
[45] Lin H. (2007) piRNAs in the germ line. Science, 316(5823)397. https://doi.org/10.1126/science.1137543
[46] Lu M, Dong X, Zhang Z, Li W, Khoshnam SE. (2020) Non-coding RNAs in Ischemic Stroke: Roles in the Neuroinflammation and Cell Death. Neurotox Res, 38(3)564-78. https://doi.org/10.1007/s12640-020-00236-7
[47] Luo M, Hao L, Hu F, Dong Y, Gou L, Zhang W, et al. (2015) MicroRNA profiles and potential regulatory pattern during the early stage of spermatogenesis in mice. Sci China Life Sci, 58(5)442-50. https://doi.org/10.1007/s11427-014-4737-8
[48] Maejima H, Okamura M, Inoue T, Takamatsu Y, Nishio T, Liu Y. (2023) Epigenetic modifications in the motor cortex caused by exercise or pharmacological inhibition of histone deacetylases (HDACs) after intracerebral hemorrhage (ICH). Brain Res, 1806148286. https://doi.org/10.1016/j.brainres.2023.148286
[49] Maes T, Mascaro C, Ortega A, Lunardi S, Ciceri F, Somervaille TC, et al. (2015) KDM1 histone lysine demethylases as targets for treatments of oncological and neurodegenerative disease. Epigenomics, 7(4)609-26. https://doi.org/10.2217/epi.15.9
[50] Mandumpala JJ, Baby S, Tom AA, Godugu C, Shankaraiah N. (2022) Role of histone demethylases and histone methyltransferases in triple-negative breast cancer: Epigenetic mnemonics. Life Sci, 292120321. https://doi.org/10.1016/j.lfs.2022.120321
[51] Mercer TR, Dinger ME, Mattick JS. (2009) Long non-coding RNAs: insights into functions. Nat Rev Genet, 10(3)155-9. https://doi.org/10.1038/nrg2521
[52] Moazed D. (2009) Small RNAs in transcriptional gene silencing and genome defence. Nature, 457(7228)413-20. https://doi.org/10.1038/nature07756
[53] Morris KV, Chan SW, Jacobsen SE, Looney DJ. (2004) Small interfering RNA-induced transcriptional gene silencing in human cells. Science, 305(5688)1289-92. https://doi.org/10.1126/science.1101372
[54] Ng GY, Lim YA, Sobey CG, Dheen T, Fann DY, Arumugam TV. (2018) Epigenetic regulation of inflammation in stroke. Ther Adv Neurol Disord, 111756286418771815. https://doi.org/10.1177/1756286418771815
[55] Ponting CP, Oliver PL, Reik W. (2009) Evolution and functions of long noncoding RNAs. Cell, 136(4)629-41. https://doi.org/10.1016/j.cell.2009.02.006
[56] Rechtsteiner A, Ercan S, Takasaki T, Phippen TM, Egelhofer TA, Wang W, et al. (2010) The histone H3K36 methyltransferase MES-4 acts epigenetically to transmit the memory of germline gene expression to progeny. PLoS Genet, 6(9)e1001091. https://doi.org/10.1371/journal.pgen.1001091
[57] Ruvkun G. (2001) Molecular biology. Glimpses of a tiny RNA world. Science, 294(5543)797-9. https://doi.org/10.1126/science.1066315
[58] Schweizer S, Meisel A, Marschenz S. (2013) Epigenetic mechanisms in cerebral ischemia. J Cereb Blood Flow Metab, 33(9)1335-46. https://doi.org/10.1038/jcbfm.2013.93
[59] Sharma AR, Shashikiran U, Uk AR, Shetty R, Satyamoorthy K, Rai PS. (2020) Aberrant DNA methylation and miRNAs in coronary artery diseases and stroke: a systematic review. Brief Funct Genomics, 19(4)259-85. https://doi.org/10.1093/bfgp/elz043
[60] Stanzione R, Cotugno M, Bianchi F, Marchitti S, Forte M, Volpe M, et al. (2020) Pathogenesis of Ischemic Stroke: Role of Epigenetic Mechanisms. Genes (Basel), 11(1). https://doi.org/10.3390/genes11010089
[61] Struhl K. (1998) Histone acetylation and transcriptional regulatory mechanisms. Genes Dev, 12(5)599-606. https://doi.org/10.1101/gad.12.5.599
[62] Tan KS, Armugam A, Sepramaniam S, Lim KY, Setyowati KD, Wang CW, et al. (2009) Expression profile of MicroRNAs in young stroke patients. PLoS One, 4(11)e7689. https://doi.org/10.1371/journal.pone.0007689
[63] Tchurikov NA. (2005) Molecular mechanisms of epigenetics. Biochemistry (Mosc), 70(4)406-23. https://doi.org/10.1007/s10541-005-0131-2
[64] Vire E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C, et al. (2006) The Polycomb group protein EZH2 directly controls DNA methylation. Nature, 439(7078)871-4. https://doi.org/10.1038/nature04431
[65] Wang KC, Chang HY. (2011) Molecular mechanisms of long noncoding RNAs. Mol Cell, 43(6)904-14. https://doi.org/10.1016/j.molcel.2011.08.018
[66] Wang SW, Liu Z, Shi ZS. (2018) Non-Coding RNA in Acute Ischemic Stroke: Mechanisms, Biomarkers and Therapeutic Targets. Cell Transplant, 27(12)1763-77. https://doi.org/10.1177/0963689718806818
[67] Wang Y, Zhang X, Liu H, Zhou X. (2021) Chemical methods and advanced sequencing technologies for deciphering mRNA modifications. Chem Soc Rev, 50(24)13481-97. https://doi.org/10.1039/d1cs00920f
[68] Wang Z, Tsai LK, Munasinghe J, Leng Y, Fessler EB, Chibane F, et al. (2012) Chronic valproate treatment enhances postischemic angiogenesis and promotes functional recovery in a rat model of ischemic stroke. Stroke, 43(9)2430-6. https://doi.org/10.1161/STROKEAHA.112.652545
[69] Wang Z, Zang C, Cui K, Schones DE, Barski A, Peng W, et al. (2009) Genome-wide mapping of HATs and HDACs reveals distinct functions in active and inactive genes. Cell, 138(5)1019-31. https://doi.org/10.1016/j.cell.2009.06.049
[70] Weake VM, Workman JL. (2008) Histone ubiquitination: triggering gene activity. Mol Cell, 29(6)653-63. https://doi.org/10.1016/j.molcel.2008.02.014
[71] Wei JW, Huang K, Yang C, Kang CS. (2017) Non-coding RNAs as regulators in epigenetics (Review). Oncol Rep, 37(1)3-9. https://doi.org/10.3892/or.2016.5236
[72] Wolf PA, Grotta JC. (2000) Cerebrovascular disease. Circulation, 102(20 Suppl 4)IV75-80. https://doi.org/10.1161/01.cir.102.suppl_4.iv-75
[73] Yu H. (2009) [Epigenetics: advances of non-coding RNAs regulation in mammalian cells]. Yi Chuan, 31(11)1077-86. https://doi.org/10.3724/sp.j.1005.2009.01077
[74] Zaratiegui M, Irvine DV, Martienssen RA. (2007) Noncoding RNAs and gene silencing. Cell, 128(4)763-76. https://doi.org/10.1016/j.cell.2007.02.016
[75] Zhan L, Chen M, Pang T, Li X, Long L, Liang D, et al. (2023) Attenuation of Piwil2 induced by hypoxic postconditioning prevents cerebral ischemic injury by inhibiting CREB2 promoter methylation. Brain Pathol, 33(1)e13109. https://doi.org/10.1111/bpa.13109
[76] Zhao LY, Song J, Liu Y, Song CX, Yi C. (2020) Mapping the epigenetic modifications of DNA and RNA. Protein Cell, 11(11)792-808. https://doi.org/10.1007/s13238-020-00733-7
[77] Zhou W, Wang J, Man WY, Zhang QW, Xu WG. (2015) siRNA silencing EZH2 reverses cisplatin-resistance of human non-small cell lung and gastric cancer cells. Asian Pac J Cancer Prev, 16(6)2425-30. https://doi.org/10.7314/apjcp.2015.16.6.2425
[78] Zhu MX, Zhao TY, Li Y. (2023) Insight into the mechanism of DNA methylation and miRNA-mRNA regulatory network in ischemic stroke. Math Biosci Eng, 20(6)10264-83. https://doi.org/10.3934/mbe.2023450
 Figures
Figures References
References Peer
Peer Information
Information[1] Abe A, Tanaka M, Yasuoka A, Saito Y, Okada S, Mishina M, et al. (2020) Changes in Whole-Blood microRNA Profiles during the Onset and Treatment Process of Cerebral Infarction: A Human Study. Int J Mol Sci, 21(9). https://doi.org/10.3390/ijms21093107
[2] Amaral PP, Dinger ME, Mercer TR, Mattick JS. (2008) The eukaryotic genome as an RNA machine. Science, 319(5871)1787-9. https://doi.org/10.1126/science.1155472
[3] Baccarelli A, Tarantini L, Wright RO, Bollati V, Litonjua AA, Zanobetti A, et al. (2010) Repetitive element DNA methylation and circulating endothelial and inflammation markers in the VA normative aging study. Epigenetics, 5(3)222-8. https://doi.org/10.4161/epi.5.3.11377
[4] Bakhit Y, Schmitt I, Hamed A, Ibrahim EAA, Mohamed IN, El-Sadig SM, et al. (2022) Methylation of alpha-synuclein in a Sudanese cohort. Parkinsonism Relat Disord, 1016-8. https://doi.org/10.1016/j.parkreldis.2022.05.009
[5] Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, et al. (2007) High-resolution profiling of histone methylations in the human genome. Cell, 129(4)823-37. https://doi.org/10.1016/j.cell.2007.05.009
[6] Basavarajappa BS, Subbanna S. (2021) Histone Methylation Regulation in Neurodegenerative Disorders. Int J Mol Sci, 22(9). https://doi.org/10.3390/ijms22094654
[7] Bayne EH, Allshire RC. (2005) RNA-directed transcriptional gene silencing in mammals. Trends Genet, 21(7)370-3. https://doi.org/10.1016/j.tig.2005.05.007
[8] Cao X, Yeo G, Muotri AR, Kuwabara T, Gage FH. (2006) Noncoding RNAs in the mammalian central nervous system. Annu Rev Neurosci, 2977-103. https://doi.org/10.1146/annurev.neuro.29.051605.112839
[9] Carthew RW, Sontheimer EJ. (2009) Origins and Mechanisms of miRNAs and siRNAs. Cell, 136(4)642-55. https://doi.org/10.1016/j.cell.2009.01.035
[10] Chen J, Liu P, Dong X, Jin J, Xu Y. (2021) The role of lncRNAs in ischemic stroke. Neurochem Int, 147105019. https://doi.org/10.1016/j.neuint.2021.105019
[11] Collaborators GBDSRF. (2024) Global, regional, and national burden of stroke and its risk factors, 1990-2021: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Neurol, 23(10)973-1003. https://doi.org/10.1016/S1474-4422(24)00369-7
[12] Costa FF. (2008) Non-coding RNAs, epigenetics and complexity. Gene, 410(1)9-17. https://doi.org/10.1016/j.gene.2007.12.008
[13] Cullell N, Soriano-Tarraga C, Gallego-Fabrega C, Carcel-Marquez J, Torres-Aguila NP, Muino E, et al. (2022) DNA Methylation and Ischemic Stroke Risk: An Epigenome-Wide Association Study. Thromb Haemost, 122(10)1767-78. https://doi.org/10.1055/s-0042-1749328
[14] Demyanenko S, Berezhnaya E, Neginskaya M, Rodkin S, Dzreyan V, Pitinova M. (2019) capital ES, Cyrilliclass II histone deacetylases in the post-stroke recovery period-expression, cellular, and subcellular localization-promising targets for neuroprotection. J Cell Biochem, 120(12)19590-609. https://doi.org/10.1002/jcb.29266
[15] Demyanenko S, Neginskaya M, Berezhnaya E. (2018) Expression of Class I Histone Deacetylases in Ipsilateral and Contralateral Hemispheres after the Focal Photothrombotic Infarction in the Mouse Brain. Transl Stroke Res, 9(5)471-83. https://doi.org/10.1007/s12975-017-0595-6
[16] Demyanenko S, Uzdensky A. (2019) Epigenetic Alterations Induced by Photothrombotic Stroke in the Rat Cerebral Cortex: Deacetylation of Histone h3, Upregulation of Histone Deacetylases and Histone Acetyltransferases. Int J Mol Sci, 20(12). https://doi.org/10.3390/ijms20122882
[17] Deng P, Chen QM, Hong C, Wang CY. (2015) Histone methyltransferases and demethylases: regulators in balancing osteogenic and adipogenic differentiation of mesenchymal stem cells. Int J Oral Sci, 7(4)197-204. https://doi.org/10.1038/ijos.2015.41
[18] Feng J, Chang H, Li E, Fan G. (2005) Dynamic expression of de novo DNA methyltransferases Dnmt3a and Dnmt3b in the central nervous system. J Neurosci Res, 79(6)734-46. https://doi.org/10.1002/jnr.20404
[19] Fujii R, Yamada H, Munetsuna E, Yamazaki M, Mizuno G, Tsuboi Y, et al. (2019) Dietary vegetable intake is inversely associated with ATP-binding cassette protein A1 (ABCA1) DNA methylation levels among Japanese women. Nutrition, 651-5. https://doi.org/10.1016/j.nut.2019.02.010
[20] Gallego-Fabrega C, Carrera C, Reny JL, Fontana P, Slowik A, Pera J, et al. (2016) TRAF3 Epigenetic Regulation Is Associated With Vascular Recurrence in Patients With Ischemic Stroke. Stroke, 47(5)1180-6. https://doi.org/10.1161/STROKEAHA.115.012237
[21] Gediya P, Parikh PK, Vyas VK, Ghate MD. (2021) Histone deacetylase 2: A potential therapeutic target for cancer and neurodegenerative disorders. Eur J Med Chem, 216113332. https://doi.org/10.1016/j.ejmech.2021.113332
[22] Ghafouri-Fard S, Shoorei H, Taheri M. (2020) Non-coding RNAs participate in the ischemia-reperfusion injury. Biomed Pharmacother, 129110419. https://doi.org/10.1016/j.biopha.2020.110419
[23] Ghildiyal M, Zamore PD. (2009) Small silencing RNAs: an expanding universe. Nat Rev Genet, 10(2)94-108. https://doi.org/10.1038/nrg2504
[24] Goto K, Numata M, Komura JI, Ono T, Bestor TH, Kondo H. (1994) Expression of DNA methyltransferase gene in mature and immature neurons as well as proliferating cells in mice. Differentiation, 56(1-2)39-44. https://doi.org/10.1046/j.1432-0436.1994.56120039.x
[25] Greer EL, Shi Y. (2012) Histone methylation: a dynamic mark in health, disease and inheritance. Nat Rev Genet, 13(5)343-57. https://doi.org/10.1038/nrg3173
[26] Griffiths-Jones S, Grocock RJ, van Dongen S, Bateman A, Enright AJ. (2006) miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res, 34(Database issue)D140-4. https://doi.org/10.1093/nar/gkj112
[27] Gurley LR, Walters RA, Tobey RA. (1973) Histone phosphorylation in late interphase and mitosis. Biochem Biophys Res Commun, 50(3)744-50. https://doi.org/10.1016/0006-291x(73)91307-7
[28] He S, Ye X, Duan R, Zhao Y, Wei Y, Wang Y, et al. (2022) Epigenome-Wide Association Study Reveals Differential Methylation Sites and Association of Gene Expression Regulation with Ischemic Moyamoya Disease in Adults. Oxid Med Cell Longev, 20227192060. https://doi.org/10.1155/2022/7192060
[29] Holliday R. (2006) Epigenetics: a historical overview. Epigenetics, 1(2)76-80. https://doi.org/10.4161/epi.1.2.2762
[30] Hu Z, Zhong B, Tan J, Chen C, Lei Q, Zeng L. (2017) The Emerging Role of Epigenetics in Cerebral Ischemia. Mol Neurobiol, 54(3)1887-905. https://doi.org/10.1007/s12035-016-9788-3
[31] Ito S, D'Alessio AC, Taranova OV, Hong K, Sowers LC, Zhang Y. (2010) Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature, 466(7310)1129-33. https://doi.org/10.1038/nature09303
[32] Jhelum P, Karisetty BC, Kumar A, Chakravarty S. (2017) Implications of Epigenetic Mechanisms and their Targets in Cerebral Ischemia Models. Curr Neuropharmacol, 15(6)815-30. https://doi.org/10.2174/1570159X14666161213143907
[33] Jiang D, Wang Y, Shen Y, Xu Y, Zhu H, Wang J, et al. (2016) Estrogen and promoter methylation in the regulation of PLA2G7 transcription. Gene, 591(1)262-7. https://doi.org/10.1016/j.gene.2016.07.048
[34] Jinek M, Doudna JA. (2009) A three-dimensional view of the molecular machinery of RNA interference. Nature, 457(7228)405-12. https://doi.org/10.1038/nature07755
[35] Joensuu EI, Nieminen TT, Lotsari JE, Pavicic W, Abdel-Rahman WM, Peltomaki P. (2015) Methyltransferase expression and tumor suppressor gene methylation in sporadic and familial colorectal cancer. Genes Chromosomes Cancer, 54(12)776-87. https://doi.org/10.1002/gcc.22289
[36] Jones PA. (2012) Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet, 13(7)484-92. https://doi.org/10.1038/nrg3230
[37] Jones PA, Baylin SB. (2007) The epigenomics of cancer. Cell, 128(4)683-92. https://doi.org/10.1016/j.cell.2007.01.029
[38] Kassis H, Shehadah A, Chopp M, Zhang ZG. (2017) Epigenetics in Stroke Recovery. Genes (Basel), 8(3). https://doi.org/10.3390/genes8030089
[39] Kawasaki H, Taira K. (2004) Induction of DNA methylation and gene silencing by short interfering RNAs in human cells. Nature, 431(7005)211-7. https://doi.org/10.1038/nature02889
[40] Kim M, Long TI, Arakawa K, Wang R, Yu MC, Laird PW. (2010) DNA methylation as a biomarker for cardiovascular disease risk. PLoS One, 5(3)e9692. https://doi.org/10.1371/journal.pone.0009692
[41] Kozomara A, Griffiths-Jones S. (2014) miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res, 42(Database issue)D68-73. https://doi.org/10.1093/nar/gkt1181
[42] Lan T, Hu L, Sun T, Wang X, Xiao Z, Shen D, et al. (2023) H3K9 trimethylation dictates neuronal ferroptosis through repressing Tfr1. J Cereb Blood Flow Metab, 43(8)1365-81. https://doi.org/10.1177/0271678X231165653
[43] Lanzillotta A, Pignataro G, Branca C, Cuomo O, Sarnico I, Benarese M, et al. (2013) Targeted acetylation of NF-kappaB/RelA and histones by epigenetic drugs reduces post-ischemic brain injury in mice with an extended therapeutic window. Neurobiol Dis, 49177-89. https://doi.org/10.1016/j.nbd.2012.08.018
[44] Li Y. (2021) Modern epigenetics methods in biological research. Methods, 187104-13. https://doi.org/10.1016/j.ymeth.2020.06.022
[45] Lin H. (2007) piRNAs in the germ line. Science, 316(5823)397. https://doi.org/10.1126/science.1137543
[46] Lu M, Dong X, Zhang Z, Li W, Khoshnam SE. (2020) Non-coding RNAs in Ischemic Stroke: Roles in the Neuroinflammation and Cell Death. Neurotox Res, 38(3)564-78. https://doi.org/10.1007/s12640-020-00236-7
[47] Luo M, Hao L, Hu F, Dong Y, Gou L, Zhang W, et al. (2015) MicroRNA profiles and potential regulatory pattern during the early stage of spermatogenesis in mice. Sci China Life Sci, 58(5)442-50. https://doi.org/10.1007/s11427-014-4737-8
[48] Maejima H, Okamura M, Inoue T, Takamatsu Y, Nishio T, Liu Y. (2023) Epigenetic modifications in the motor cortex caused by exercise or pharmacological inhibition of histone deacetylases (HDACs) after intracerebral hemorrhage (ICH). Brain Res, 1806148286. https://doi.org/10.1016/j.brainres.2023.148286
[49] Maes T, Mascaro C, Ortega A, Lunardi S, Ciceri F, Somervaille TC, et al. (2015) KDM1 histone lysine demethylases as targets for treatments of oncological and neurodegenerative disease. Epigenomics, 7(4)609-26. https://doi.org/10.2217/epi.15.9
[50] Mandumpala JJ, Baby S, Tom AA, Godugu C, Shankaraiah N. (2022) Role of histone demethylases and histone methyltransferases in triple-negative breast cancer: Epigenetic mnemonics. Life Sci, 292120321. https://doi.org/10.1016/j.lfs.2022.120321
[51] Mercer TR, Dinger ME, Mattick JS. (2009) Long non-coding RNAs: insights into functions. Nat Rev Genet, 10(3)155-9. https://doi.org/10.1038/nrg2521
[52] Moazed D. (2009) Small RNAs in transcriptional gene silencing and genome defence. Nature, 457(7228)413-20. https://doi.org/10.1038/nature07756
[53] Morris KV, Chan SW, Jacobsen SE, Looney DJ. (2004) Small interfering RNA-induced transcriptional gene silencing in human cells. Science, 305(5688)1289-92. https://doi.org/10.1126/science.1101372
[54] Ng GY, Lim YA, Sobey CG, Dheen T, Fann DY, Arumugam TV. (2018) Epigenetic regulation of inflammation in stroke. Ther Adv Neurol Disord, 111756286418771815. https://doi.org/10.1177/1756286418771815
[55] Ponting CP, Oliver PL, Reik W. (2009) Evolution and functions of long noncoding RNAs. Cell, 136(4)629-41. https://doi.org/10.1016/j.cell.2009.02.006
[56] Rechtsteiner A, Ercan S, Takasaki T, Phippen TM, Egelhofer TA, Wang W, et al. (2010) The histone H3K36 methyltransferase MES-4 acts epigenetically to transmit the memory of germline gene expression to progeny. PLoS Genet, 6(9)e1001091. https://doi.org/10.1371/journal.pgen.1001091
[57] Ruvkun G. (2001) Molecular biology. Glimpses of a tiny RNA world. Science, 294(5543)797-9. https://doi.org/10.1126/science.1066315
[58] Schweizer S, Meisel A, Marschenz S. (2013) Epigenetic mechanisms in cerebral ischemia. J Cereb Blood Flow Metab, 33(9)1335-46. https://doi.org/10.1038/jcbfm.2013.93
[59] Sharma AR, Shashikiran U, Uk AR, Shetty R, Satyamoorthy K, Rai PS. (2020) Aberrant DNA methylation and miRNAs in coronary artery diseases and stroke: a systematic review. Brief Funct Genomics, 19(4)259-85. https://doi.org/10.1093/bfgp/elz043
[60] Stanzione R, Cotugno M, Bianchi F, Marchitti S, Forte M, Volpe M, et al. (2020) Pathogenesis of Ischemic Stroke: Role of Epigenetic Mechanisms. Genes (Basel), 11(1). https://doi.org/10.3390/genes11010089
[61] Struhl K. (1998) Histone acetylation and transcriptional regulatory mechanisms. Genes Dev, 12(5)599-606. https://doi.org/10.1101/gad.12.5.599
[62] Tan KS, Armugam A, Sepramaniam S, Lim KY, Setyowati KD, Wang CW, et al. (2009) Expression profile of MicroRNAs in young stroke patients. PLoS One, 4(11)e7689. https://doi.org/10.1371/journal.pone.0007689
[63] Tchurikov NA. (2005) Molecular mechanisms of epigenetics. Biochemistry (Mosc), 70(4)406-23. https://doi.org/10.1007/s10541-005-0131-2
[64] Vire E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C, et al. (2006) The Polycomb group protein EZH2 directly controls DNA methylation. Nature, 439(7078)871-4. https://doi.org/10.1038/nature04431
[65] Wang KC, Chang HY. (2011) Molecular mechanisms of long noncoding RNAs. Mol Cell, 43(6)904-14. https://doi.org/10.1016/j.molcel.2011.08.018
[66] Wang SW, Liu Z, Shi ZS. (2018) Non-Coding RNA in Acute Ischemic Stroke: Mechanisms, Biomarkers and Therapeutic Targets. Cell Transplant, 27(12)1763-77. https://doi.org/10.1177/0963689718806818
[67] Wang Y, Zhang X, Liu H, Zhou X. (2021) Chemical methods and advanced sequencing technologies for deciphering mRNA modifications. Chem Soc Rev, 50(24)13481-97. https://doi.org/10.1039/d1cs00920f
[68] Wang Z, Tsai LK, Munasinghe J, Leng Y, Fessler EB, Chibane F, et al. (2012) Chronic valproate treatment enhances postischemic angiogenesis and promotes functional recovery in a rat model of ischemic stroke. Stroke, 43(9)2430-6. https://doi.org/10.1161/STROKEAHA.112.652545
[69] Wang Z, Zang C, Cui K, Schones DE, Barski A, Peng W, et al. (2009) Genome-wide mapping of HATs and HDACs reveals distinct functions in active and inactive genes. Cell, 138(5)1019-31. https://doi.org/10.1016/j.cell.2009.06.049
[70] Weake VM, Workman JL. (2008) Histone ubiquitination: triggering gene activity. Mol Cell, 29(6)653-63. https://doi.org/10.1016/j.molcel.2008.02.014
[71] Wei JW, Huang K, Yang C, Kang CS. (2017) Non-coding RNAs as regulators in epigenetics (Review). Oncol Rep, 37(1)3-9. https://doi.org/10.3892/or.2016.5236
[72] Wolf PA, Grotta JC. (2000) Cerebrovascular disease. Circulation, 102(20 Suppl 4)IV75-80. https://doi.org/10.1161/01.cir.102.suppl_4.iv-75
[73] Yu H. (2009) [Epigenetics: advances of non-coding RNAs regulation in mammalian cells]. Yi Chuan, 31(11)1077-86. https://doi.org/10.3724/sp.j.1005.2009.01077
[74] Zaratiegui M, Irvine DV, Martienssen RA. (2007) Noncoding RNAs and gene silencing. Cell, 128(4)763-76. https://doi.org/10.1016/j.cell.2007.02.016
[75] Zhan L, Chen M, Pang T, Li X, Long L, Liang D, et al. (2023) Attenuation of Piwil2 induced by hypoxic postconditioning prevents cerebral ischemic injury by inhibiting CREB2 promoter methylation. Brain Pathol, 33(1)e13109. https://doi.org/10.1111/bpa.13109
[76] Zhao LY, Song J, Liu Y, Song CX, Yi C. (2020) Mapping the epigenetic modifications of DNA and RNA. Protein Cell, 11(11)792-808. https://doi.org/10.1007/s13238-020-00733-7
[77] Zhou W, Wang J, Man WY, Zhang QW, Xu WG. (2015) siRNA silencing EZH2 reverses cisplatin-resistance of human non-small cell lung and gastric cancer cells. Asian Pac J Cancer Prev, 16(6)2425-30. https://doi.org/10.7314/apjcp.2015.16.6.2425
[78] Zhu MX, Zhao TY, Li Y. (2023) Insight into the mechanism of DNA methylation and miRNA-mRNA regulatory network in ischemic stroke. Math Biosci Eng, 20(6)10264-83. https://doi.org/10.3934/mbe.2023450
Peer-review Terminology
Identity transparency: Single anonymized
Reviewer interacts with: Editor
Details
This is an open access article under the terms of the Creative Commons Attribution License(http://creativecommons.org/licenses/by/4.0/), which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Publication History
Received 2025-01-05
Accepted 2025-03-20
Published 2025-03-30