




Abstract
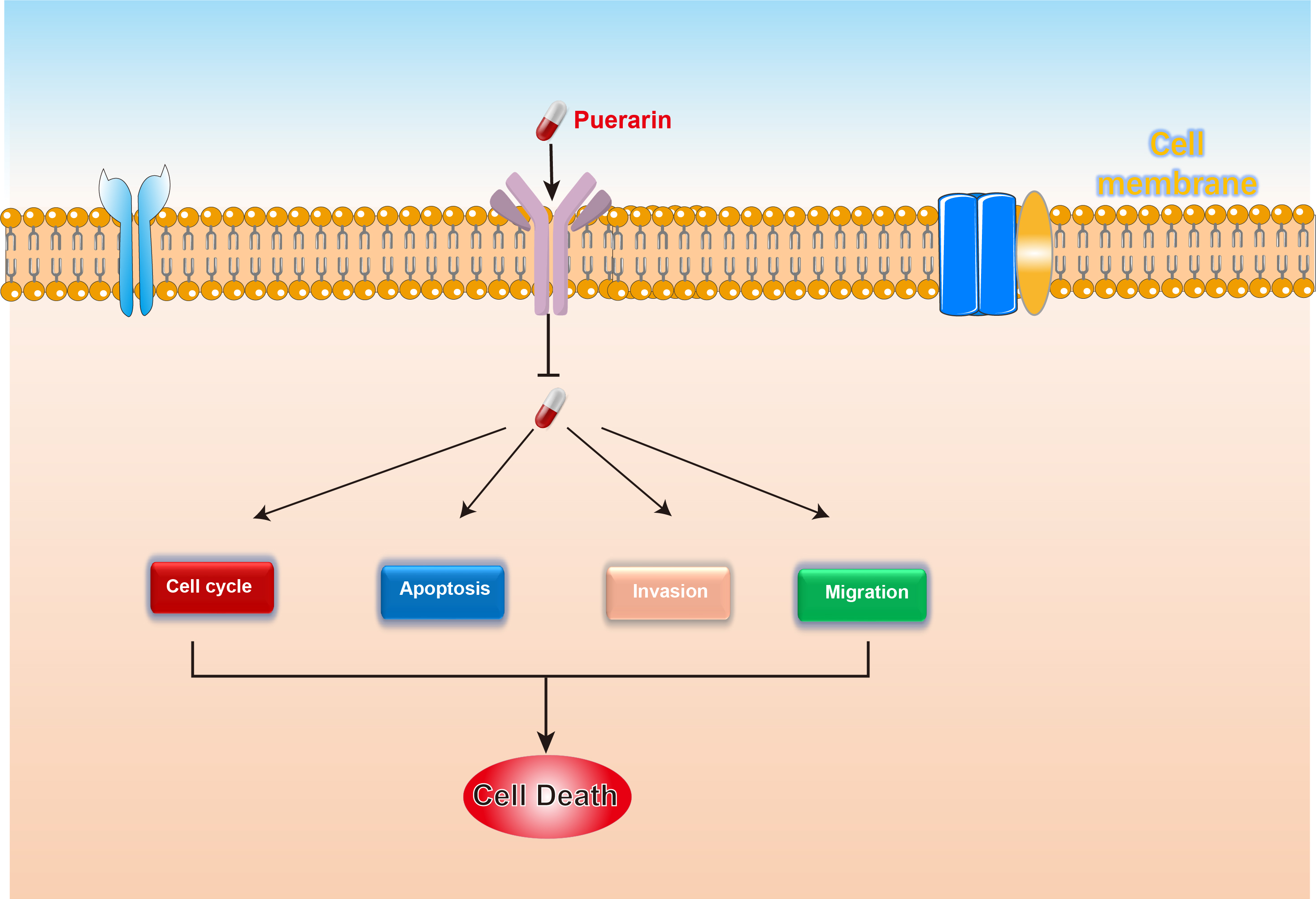
Background:Gastric cancer is a tumor with high morbidity and mortality within the world, and according to the statistics of the World Health Organization (WHO), the incidence of gastric cancer is predominant in Asia, especially in East Asia and Africa, and China and Japan are the high incidence areas of gastric cancer, which is related to the special dietary habits of East Asians.
Methods: in this study, we first investigated the effects of puerarin on the biological behaviors of two gastric cancer cell lines, HGC27 and AGS. To further explore the underlying mechanisms, we performed transcriptome sequencing, which revealed that puerarin primarily influences gastric cancer progression by regulating cell cycle transitions. To validate these findings, we examined the expression levels of cell cycle-related proteins and analyzed cell cycle distribution using flow cytometry.
Results: This study demonstrated that puerarin significantly inhibits the proliferation, migration, and invasion of gastric cancer cells (HGC27 and AGS) while promoting their apoptosis. Transcriptome sequencing analysis revealed that puerarin primarily affects the biological behavior of gastric cancer cells by regulating the G2-to-M phase transition. To validate this mechanism, we further employed flow cytometry to assess cell cycle distribution and analyzed the expression levels of cell cycle-related proteins, providing proteomic evidence that supports the G2/M phase transition regulation identified by transcriptomic data.
Conclusion:Puerarin effectively inhibits the proliferation and invasion of gastric cancer cells, and its mechanism is closely related to the regulation of the G2-to-M phase transition, thereby affecting cell cycle progression and proliferative capacity.
Keywords: Puerarin; Gastric cancer; Cell cycle
Introduction
Cancer is the second leading cause of death in the world, and gastric cancer has become an important health problem worldwide due to its high mortality rate[1]. Gastric cancer has a complex pathological mechanism, with a high degree of cellular heterogeneity[2] and a lack of specific clinical symptoms in the early stages of the disease, which leads to the majority of patients being in the middle and late stages when diagnosed and missing the best time for treatment. This is one of the main reasons for the high mortality rate[3]. The pathogenesis of gastric cancer involves the interaction of multiple factors, mainly including infectious factors and dietary factors. Among the infectious factors, Helicobacter pylori infection is considered to be the most important risk factor for gastric cancer, which may ultimately lead to gastric cancer by triggering chronic gastritis, gastric mucosal atrophy, and intestinal epithelial metaplasia[4]. In addition, Epstein-Barr virus (EBV) infection is also closely related to some gastric cancers (especially lymphoepithelioma-like gastric cancer). Among dietary factors, high salt diet and excessive intake of red meat and processed meat are important triggers of gastric cancer[5]. High salt diet not only directly damages the gastric mucosa, but also promotes the conversion of nitrite into the potent carcinogen nitrosamine in the stomach, and certain ingredients in red meat and processed meat may also increase the risk of gastric cancer[6]. Currently, the treatment of gastric cancer is relatively homogeneous, relying mainly on surgical resection, chemotherapy, and radiotherapy. For early gastric cancer, surgical resection is the main method of eradication, but for patients with intermediate and advanced stages, the therapeutic effect is often limited[7]. In addition, with the emergence of drug-resistant strains, the efficacy of chemotherapy and radiotherapy gradually decrease, which has become a major problem in gastric cancer treatment. In recent years, the rise of targeted therapy and immunotherapy has provided a new direction for the treatment of gastric cancer[8,9]. For example, for patients with HER2-positive gastric cancer, targeted drugs such as trastuzumab have shown certain efficacy. The cell cycle is the entire process that a cell undergoes from the end of one division to the end of the next, and is the central biological mechanism of cell proliferation[10].The cell cycle is divided into four main phases: G1 phase (preparation for cell growth and metabolism), S phase (DNA replication), G2 phase (preparation for mitosis) and M phase (mitosis). Normally, the cell cycle is tightly regulated to ensure that cell proliferation matches the needs of the organism and maintains tissue homeostasis. However, in tumour cells, the regulatory mechanisms of the cell cycle are disrupted, leading to uncontrolled cell proliferation, which is one of the key features of tumourigenesis and progression[11,12]. The normal transformation of the cell cycle is largely dependent on the co-regulation between cytosolic proteins and cytosolic protein-dependent kinases[13]. Cyclins form complexes with CDKs and phosphorylate downstream target proteins to drive cell cycle progression. In addition, cell cycle protein-dependent kinase inhibitors (e.g., p21, p27) and tumour suppressor proteins (e.g., p53, Rb) play key roles in cell cycle checkpoints to ensure that cells move to the next phase with DNA integrity and replication complete[14,15]. In the cell cycle, the G1/S checkpoint and the G2/M checkpoint are two important regulatory nodes[16]. At the G1/S checkpoint, the cell detects whether DNA is intact and growth signals are appropriate to decide whether to enter S phase. p53 and Rb proteins play key roles in this checkpoint. At the G2/M checkpoint, the cell detects whether DNA replication is complete and ensures that it is suitable for mitosis[17]. If this checkpoint is out of control, the cell may enter division with incompletely replicated DNA, further exacerbating genomic instability. In most tumours, abnormalities in cell cycle regulatory mechanisms are central to the excessive proliferation of tumour cells. For example, overexpression of molecules such as Cyclin D1, Cyclin E, and CDK4/6, as well as inactivation of CDKIs (e.g., p21, p27), leads to accelerated cell cycle progression. With the in-depth study of cell cycle regulatory mechanisms, studies have found that the cell cycle is closely related to tumour therapy. For example, CDK4/6 inhibitors (e.g., pabocinib, repocinib) have been approved for the treatment of tumours such as breast cancer and inhibit the proliferation of tumour cells by inhibiting the activity of the Cyclin D-CDK4/6 complex, which prevents the cell cycle from moving from the G1 phase to the S phase[18]. In addition, in the process of radiotherapy and chemotherapy, ionising radiation and chemotherapeutic drugs (e.g., cisplatin, paclitaxel) are effective in the treatment of tumours by inducing DNA damage and activating cell cycle checkpoints, leading to cell cycle arrest or apoptosis, thus inhibiting the proliferation of tumour cells[11]. Puerarin is an isoflavonoid compound extracted from the traditional Chinese medicine pueraria lobata with a wide range of biological activities and pharmacological effects[19,20]. Puerarin has a wide range of pharmacological effects, including cardiovascular protection (e.g., vasodilation, improved microcirculation, lowered blood pressure, anti-myocardial ischemia, and anti-arrhythmia), antioxidant effects (e.g., scavenging free radicals...), anti-inflammatory effects (e.g., inhibiting the release of inflammatory mediators, reducing inflammatory response); neuroprotection (e.g., improving cerebral ischemia and brain damage, protecting neurons, and slowing down the progress of neurodegenerative diseases (e.g., Alzheimer's disease, Parkinson's disease))[21,22]. It also exhibits anti-tumour effects by inhibiting proliferation of tumour cells, inducing apoptosis of tumour cells, and inhibiting tumour angiogenesis. In breast cancer, puerarin inhibits the proliferation and metastasis of breast cancer cells by inhibiting the estrogen receptor (ER) signalling pathway[23]. In hepatocellular carcinoma, it significantly inhibits the growth of hepatocellular carcinoma by inducing apoptosis and inhibiting angiogenesis[24]. In lung cancer, puerarin inhibits the proliferation and invasion of lung cancer cells through the modulation of the PI3K/Akt and MAPK signalling pathways[25]. The present study mainly explored the effect of puerarin on gastric cancer cells, and it was found that puerarin affects the proliferation of gastric cancer cells mainly by influencing the cell cycle progression. In this study, we first systematically investigated the effects of puerarin on the biological functions of gastric cancer cells. The experimental results showed that puerarin markedly suppressed the proliferation and invasion ability of gastric cancer cells, and at the same time induced tumour cell apoptosis. In order to further reveal the mechanism of action of puerarin against gastric cancer, we carried out an in-depth analysis of its molecular mechanism by using transcriptome sequencing technology. The results showed that puerarin inhibited the proliferation of gastric cancer cells mainly by regulating the expression of cell cycle-related genes and blocking the cell cycle process. To verify the anti-tumour effect of puerarin in vivo, we constructed a subcutaneous transplantation tumour model of gastric cancer to simulate the tumour microenvironment in vivo. The results of animal experiments showed that puerarin was able to significantly inhibit the subcutaneous tumour-forming ability of gastric cancer cells and reduced the tumour volume and weight, with no obvious toxic side effects observed. This finding not only confirmed the anti-tumour activity of puerarin in vivo, but also provided an important experimental basis for its potential use as a drug for gastric cancer treatment.
Materials and methods
Western blot
Proteins in tissues or cells were extracted with RIPA lysate (Beyotime, Shanghai, China) with
the addition of PMSF and phosphatase inhibitors. Then proteins were separated by SDS-PAG, transferred using PVDF
membranes, and blotted with suitable primary antibodies. The primary antibodies used were: GAPDH (Proteintech Group,
Wuhan, China; dilution 1:5000), CDK1, CDK4, Cyclin A, Cyclin B, BAX, and BCL2 (all from Cell Signaling Technology,
Shanghai, China; dilution 1:1000).
Transwell and wound-healing assays
Transwell inserts (Corning, NY, USA), either coated or uncoated with BD
Matrigel, were used to assess the invasive or migratory capacity of the cells. A total of 5 × 10⁴ cells were
resuspended in 200 μl of serum‑free medium and loaded into the upper chamber. After cells that had successfully
invaded through the Matrigel (for invasion assays) or migrated through the membrane (for migration assays) reached the
lower surface of the membrane, they were fixed with methanol and stained with crystal violet. Images were captured
using an inverted phase‑contrast microscope (Olympus, Tokyo, Japan), and the number of stained cells was quantified by
counting them with ImageJ software.
Flow cytometry for apoptosis analysis
Apoptosis was assessed in the DMSO and dosing groups using an
apoptosis kit (Annexin V-AF647/PI Apoptosis Kit, GOONOE) and data were collected using BD FACScelesta 3 (NJ, USA). The
final results were analysed using Flowjo.
Flow cytometry for cell cycle analysis
Cell cycle was assessed in the DMSO and dosing groups using the
Cell Cycle Assay Kit GOONOE) and data were collected using BD FACScelesta 3 (NJ, USA). Final results were analysed
using Flowjo.
Animal studies
Twelve 3-4 weeks old female BALB/c thymus-free nude mice (strain: BALB/c-nu/nu) weighing
18-22 g were purchased from Jiangsu Jicui Pharmacology Co., Ltd. They were housed in SPF-grade barriers with a
temperature of 22±2°C, a relative humidity of 50±10%, a 12-h light/dark cycle, and an ad libitum intake of irradiated
sterilized chow and autoclaved water. The mice were domesticated for 7 days after purchase, and the experiments were
started after daily observation of their health condition and confirmation of no abnormality. Inoculation method: Mice
were injected with 100 μL of cell suspension subcutaneously in the right axilla (the skin was sterilized with 75%
ethanol before injection). Experimental groups (n=6/group): Control group (DMSO): 100 μL saline + 0.1% DMSO (solvent
control); Low dose group: puerarin 20 μg (dissolved in saline); High dose group: puerarin 40 μg (dissolved in saline).
Euthanasia and sample collection (21 days after inoculation): Anesthesia: Intraperitoneal injection of 5% isoflurane
(dosage: 0.1 mL/10 g) to confirm the absence of nociceptive reflexes in the mice. Cervical dislocation was performed:
the head and tail were quickly immobilized and the cervical vertebrae were pulled back to ensure instantaneous death.
Sample processing: complete excision of the tumor, weighing and portioning. Portions were placed in 4%
paraformaldehyde for fixation (stained with HE and TUNEL). This study was approved by the Animal Ethics Committee of
Anhui Medical University (Protocol No. 2024123)
RNA sequencing
RNA sequencing was performed by Genedenovo Biotechnology Ltd (Guangzhou, China) using
libraries sequenced on the Illumina sequencing platform. The samples were divided into two groups. After treating
cells with DMSO or the IC50 concentration of puerarin, we extracted RNA and sent it for sequencing.
Results
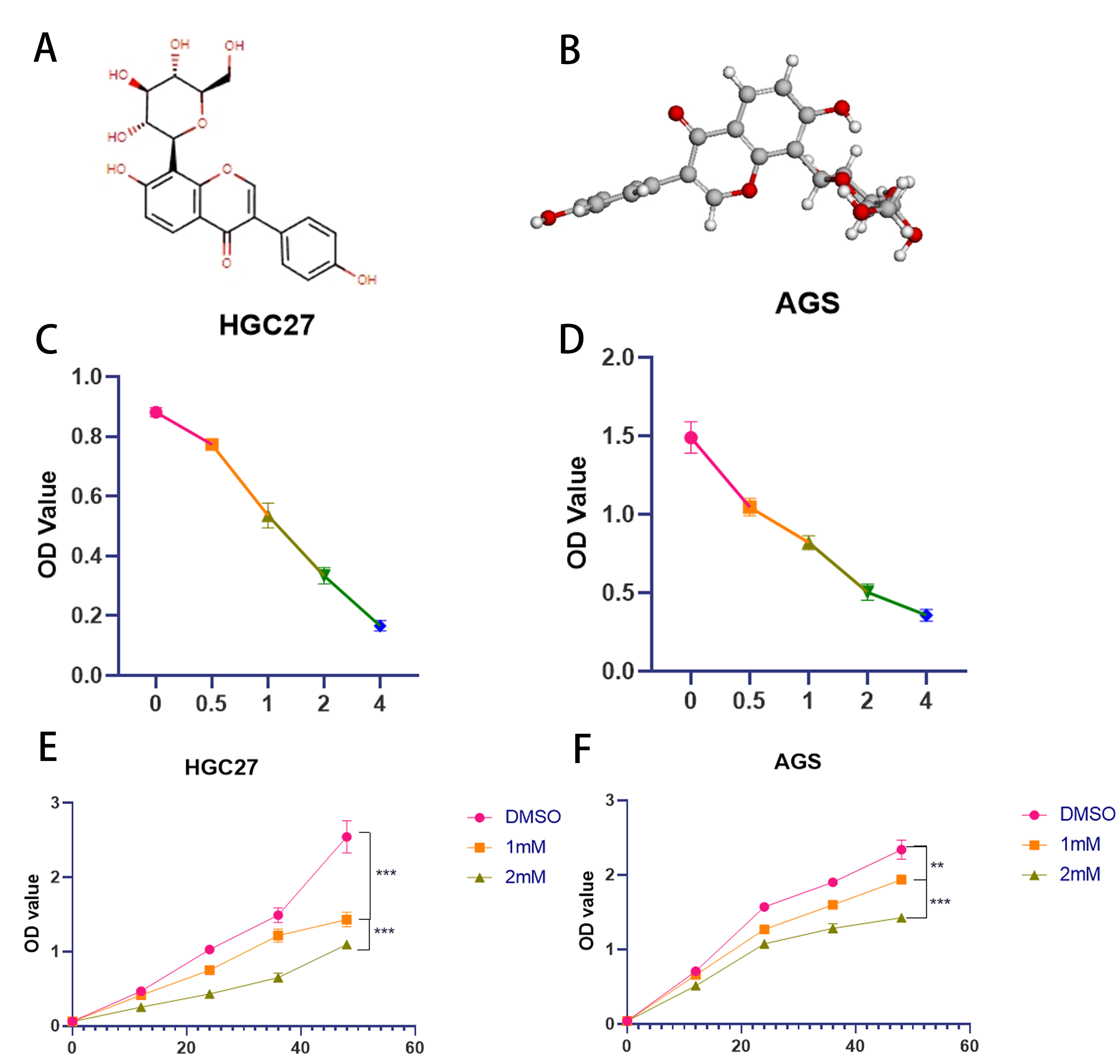
Chemical structure of puerarin and its effect on gastric cancer cell activity
The chemical structure and
three-dimensional molecular structure of puerarin are shown in Figure 1A and B. To investigate the
toxic effects of puerarin on gastric cancer cells, two commonly used gastric cancer cell lines, AGS and HGC27, were
selected as the experimental models in this study. By setting up different concentration gradients of puerarin
treatment, we determined that its optimal administration concentration for both cell lines was 1 mM (Figure 1C and D). To further investigate the effects of puerarin on gastric cancer cells, the inhibitory effects of different
concentrations of puerarin on cell proliferation over time were systematically detected using the CCK-8 (Cell Counting
Kit-8) method (Figure 1E). This experiment was designed to comprehensively evaluate the toxic
effects of puerarin on gastric cancer cells and its concentration dependence.
Figure 1. Spatial structure of Puerarin and toxic effects of two gastric cancer cells (A)Two-dimensional chemical structure of puerarin. (B) Three-dimensional molecular structure of puerarin. (C) The effect of different concentration gradients of puerarin treatment on the viability of HGC27 cells over 24 h was detected by CCK-8 method. (D) The effect of different concentration gradients of puerarin treatment on the viability of AGS cells over 24 h was detected by CCK-8 method. (E) The concentrations of DMSO (control), 1 mM, and 2 mM were used to treat HGC27 gastric cancer cells, respectively. of Puerarin were treated with HGC27 gastric cancer cells and cell viability was determined by CCK-8 assay at 0, 12, 24, 36 and 48 h time points. (F) AGS gastric cancer cells were treated with DMSO (control), 1 mM and 2 mM concentrations of Puerarin, respectively, and cell viability was determined by CCK-8 assay at 0, 12, 24, 36 and 48 h time points.
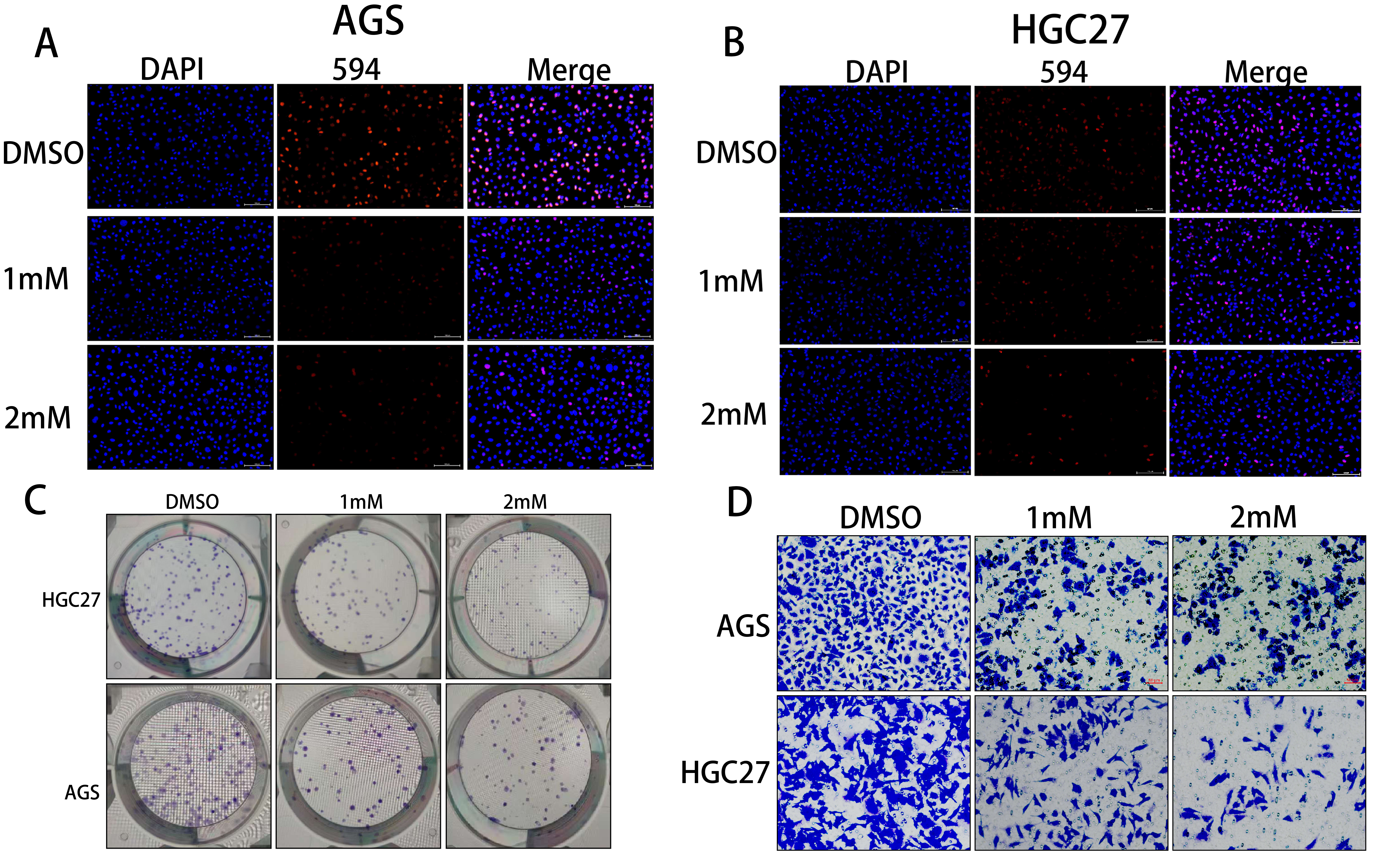
Puerarin inhibited the proliferation and invasion of gastric cancer cells.
To investigate the effect of
puerarin on the proliferation function of gastric cancer cells, The EdU and colony formation assays were performed in
this study. The results of EdU assay showed that the fluorescence positivity rate of 1 mM and 2 mM puerarin-treated
groups was significantly reduced compared with that of the DMSO control group, which indicated that puerarin could
inhibit the DNA replication activity of gastric cancer cells effectively (Figure 2A and B). The
results of colony formation assay further showed that the number of clone-forming clusters in the 1 mM and 2 mM
puerarin-treated groups was significantly reduced, which confirmed the inhibitory effect of puerarin on cell
proliferation (Figure 2C). Taken together, the above results indicated that puerarin was able to
significantly inhibit the proliferative ability of gastric cancer cells. In addition, to assess the effect of puerarin
on the invasive ability of gastric cancer cells, we used the Transwell assay. The results showed that the number of
cells passing through the lower chamber was significantly reduced in the 1 mM and 2 mM puerarin treatment groups,
indicating that puerarin was able to effectively inhibit the invasive ability of gastric cancer cells (Figure 2D).
Figure 2. Puerarin inhibited the proliferation and invasion of gastric cancer (A)The fluorescence positivity rate of AGS cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was detected by EdU assay to evaluate the effect of the drug on the proliferative viability of the cells. (B) The fluorescence positivity rate of HGC27 cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was detected by EdU assay to further evaluate the effect of Puerarin on the proliferative ability of the cells. (C) The monoclonal formation ability of AGS and HGC27 cells under different concentrations of puerarin treatment was analysed by colony formation assay to clarify the dose-dependent effect of the drug on the cell proliferation function. (D) The effects of different concentrations of puerarin treatment on the invasion ability of AGS and HGC27 cells were investigated by using Transwell assay to reveal the potential mechanism of the drug's action in inhibiting cell migration and invasion. (E) The effects of different concentrations of puerarin treatment on the invasion ability of AGS and HGC27 cells were investigated by using Transwell assay. potential mechanism of action in inhibiting cell migration and invasion.
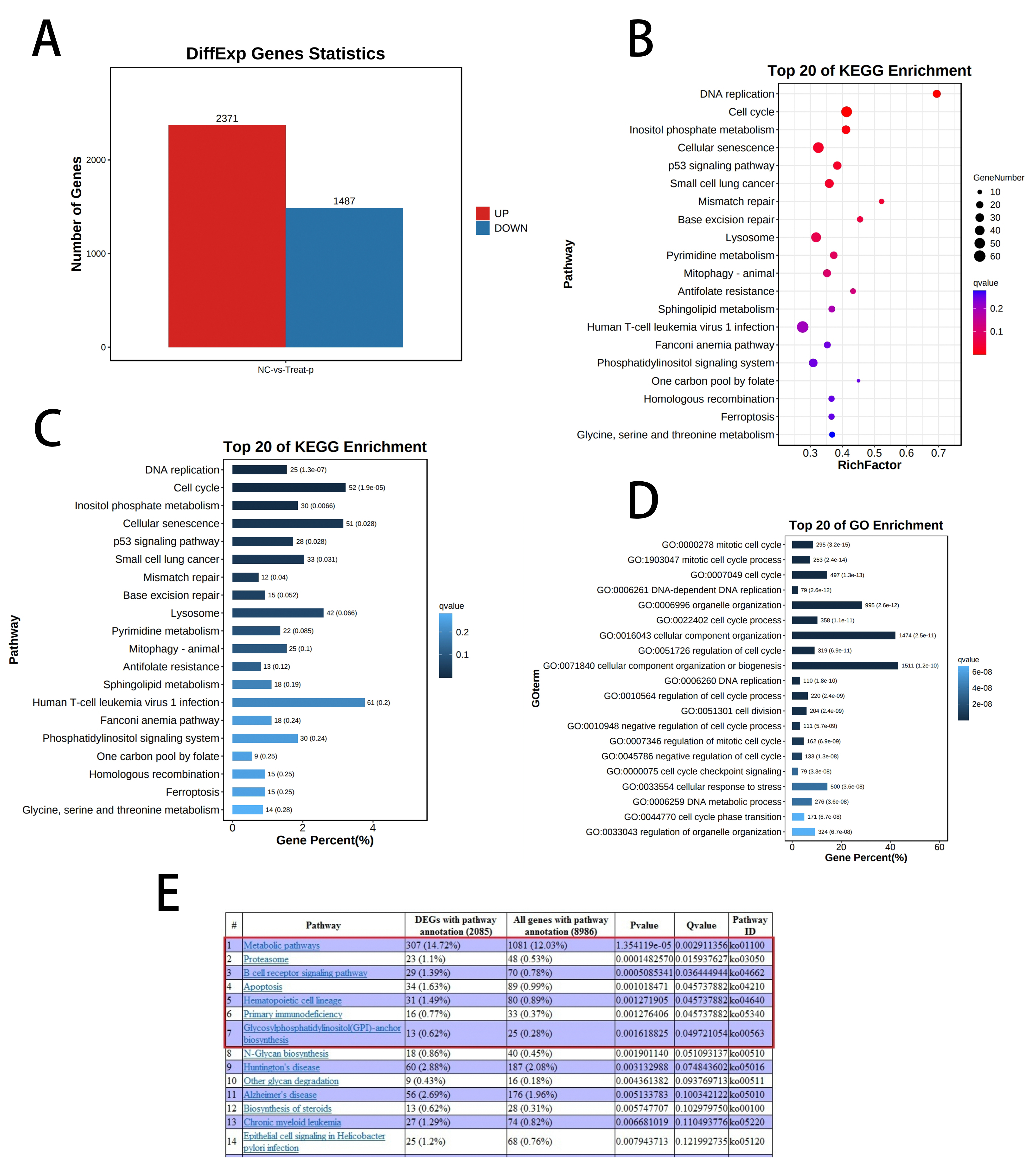
Transcriptome sequencing showed that puerarin affects tumour cell cycle progression.
In order to deeply
investigate the mechanism of the effect of puerarin on the biological behaviour of gastric cancer cells, we performed
transcriptome sequencing analysis on the DMSO control group and the 1 mM puerarin-treated group, respectively. The
sequencing results showed that there were significantly differentially expressed genes between the two groups of
samples, and a total of more than 3,000 differentially expressed genes were screened (Figure 3A). To
further elucidate the functions of these differential genes, we performed bioinformatics analyses, including KEGG
pathway enrichment analysis and GO functional annotation. The results of the analyses indicated that puerarin affected
the biological behaviours of gastric cancer cells mainly by regulating cell cycle-related pathways and DNA replication
processes (Figure 3B-D). Specifically, puerarin significantly interfered with the normal progression
of the cell cycle, leading to cell cycle arrest. In addition, puerarin was found to induce apoptosis in gastric cancer
cells, thereby effectively inhibiting the proliferation of tumour cells (Figure 3E). These findings
provide a basis for the molecular mechanism for the potential application of puerarin in gastric cancer treatment.
Figure 3. Analysis of the transcriptome sequencing results revealed the significant effects of Puerarin on the gene expression profiles of gastric cancer cells. (A) RNA sequencing results showed that more than 2,000 significantly up-regulated genes and more than 1,000 significantly down-regulated genes existed in the Puerarin-treated group compared with the control group. (B) KEGG dot plot enrichment analysis further demonstrated that the differentially expressed genes were significantly enriched in the DNA replication and cell-cycle related pathways. (C) KEGG histogram enrichment analysis results were consistent with the dot plot, which further verified that DNA replication and cell cycle pathways in the mechanism of action of puerarin. (D) GO functional annotation histogram showed that the differentially expressed genes were significantly enriched in cell cycle-related functional categories, which further supported the regulatory effect of puerarin on cell cycle progression. (E) In addition, the sequencing results also showed that puerarin significantly affected the apoptosis-related pathway, suggesting that it may inhibit the growth of gastric cancer cells by inducing cell apoptosis. growth of gastric cancer cells by inducing apoptosis.
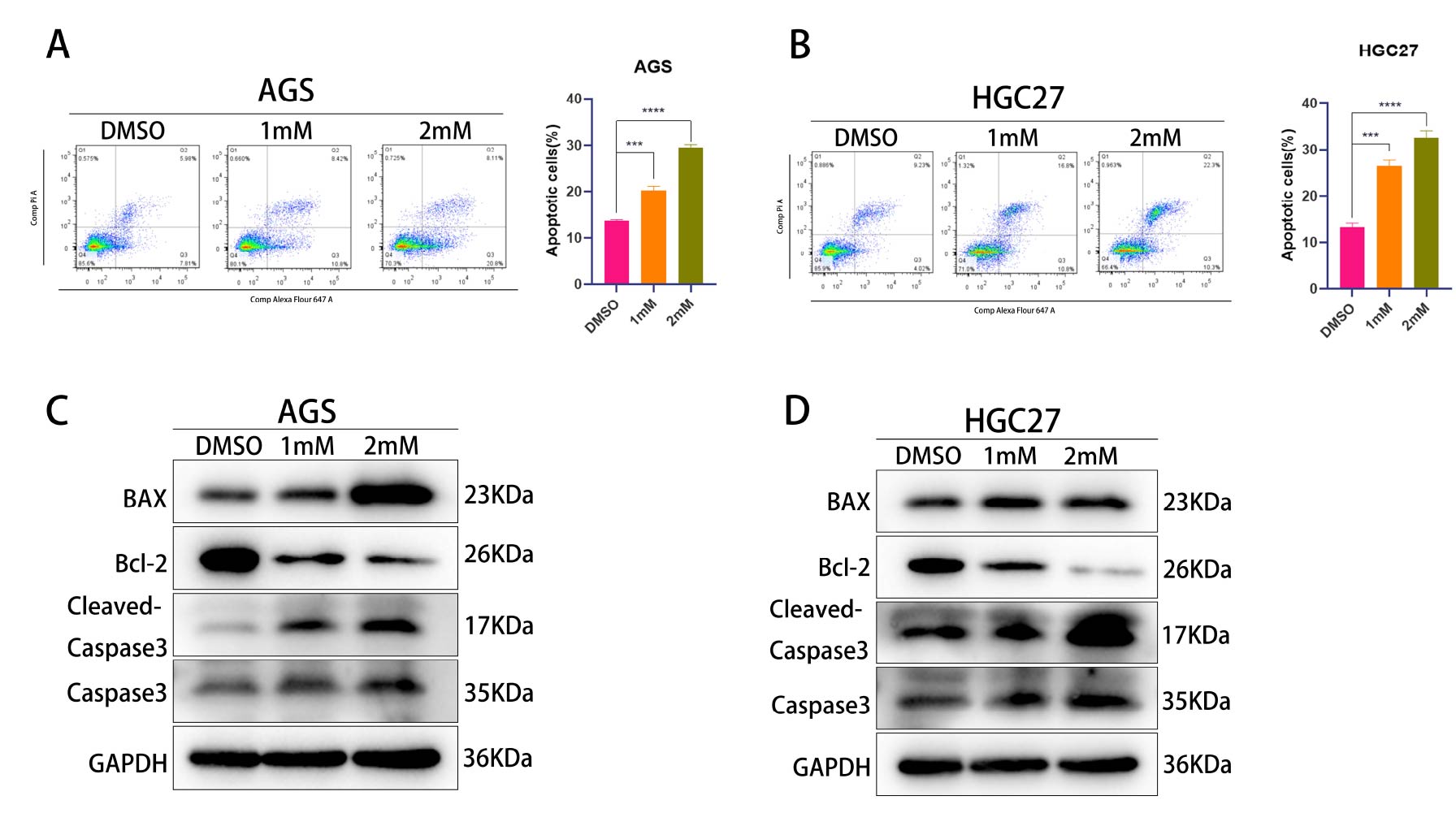
Puerarin promotes apoptosis
Sequencing results indicated that Puerarin may induce cells to undergo
apoptosis. To verify this result, we first treated AGS and HGC27 cells with different concentrations of puerarin and
detected the apoptosis rate by flow cytometry (Figure 4A and B). The results showed that the
apoptosis rate was significantly increased in the drug-treated group compared with the DMSO control group, which
initially confirmed the pro-apoptotic effect of puerarin. To further validate this phenomenon, we examined the
expression levels of apoptosis-related proteins (BAX, BCL2, Caspase3) using the Western blot technique (Figure 4C and D). The experimental results showed that puerarin treatment significantly up-regulated the expression of
pro-apoptotic proteins BAX and Caspase3, while down-regulated the expression of the anti-apoptotic protein BCL2, a
finding that was highly consistent with those detected by flow cytometry. In summary, the above experimental data
together confirmed that puerarin is able to promote apoptosis in gastric cancer cells by regulating the expression of
apoptosis-related proteins.
Figure 4. Detection of apoptosis by flow and WB experiments (A)Detection of apoptosis of AGS gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups by flow cytometry. (B) Detection of apoptosis of HGC27 gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups by flow cytometry. (C) Detection of apoptosis in DMSO (control group), 1 mM and 2 mM expression levels of AGS apoptosis-related proteins (BAX, BCL2, Caspase3) and the internal reference protein GAPDH in the Puerarin-treated groups. (D) The expression levels of apoptosis-related proteins (BAX, BCL2, Caspase3) and the internal reference protein GAPDH in HGC27 cells in the DMSO (control group), the 1 mM and the 2 mM Puerarin-treated groups were examined by Western blotting technique. protein GAPDH in the DMSO (control) and 2 mM Puerarin-treated groups
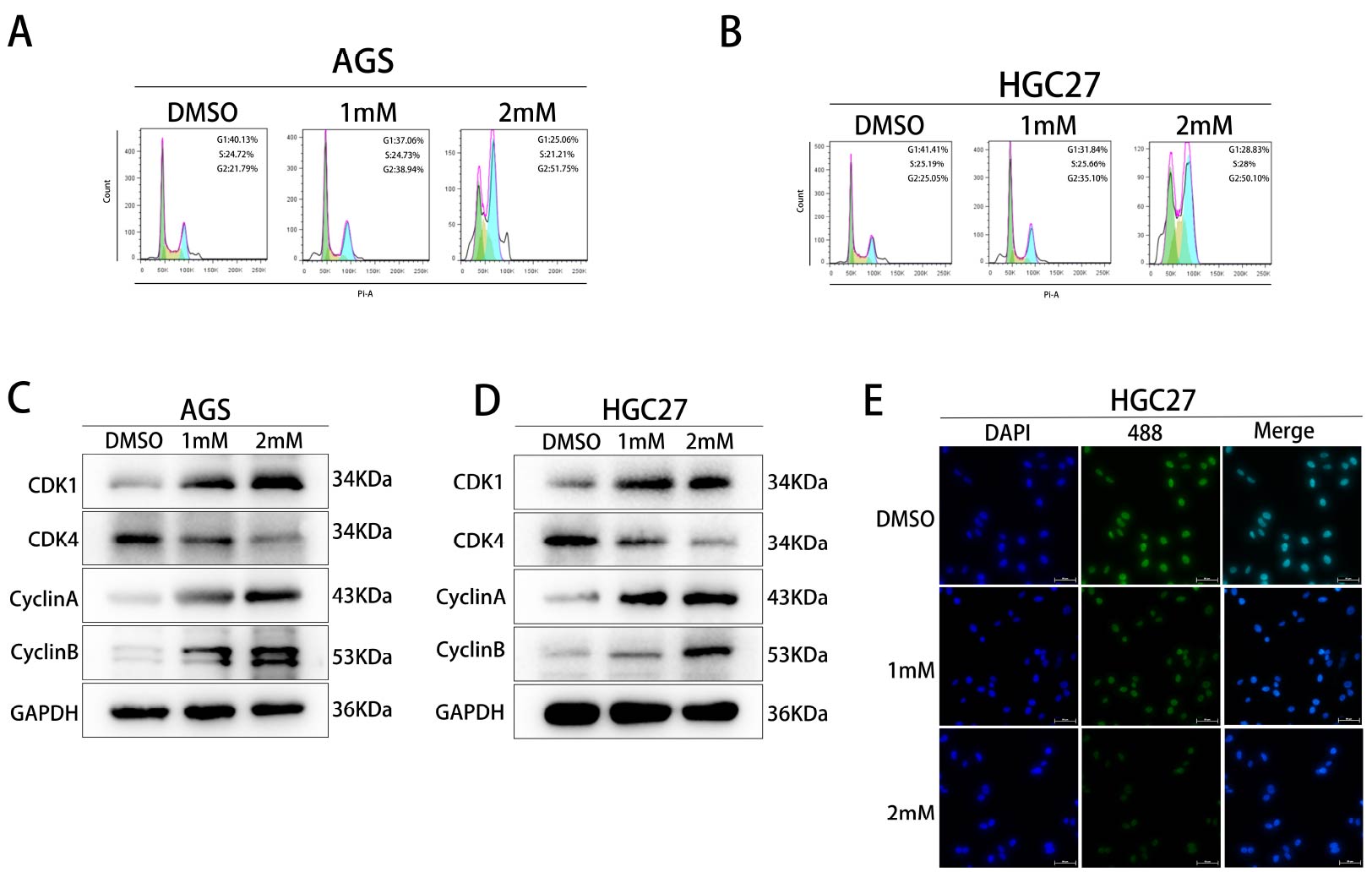
Puerarin inhibits the transition from G2 to M phase in cells.
Sequencing results indicated that puerarin
may affect the cell cycle progression of gastric cancer cells. To verify this result, we firstly treated different
concentrations of puerarin in AGS and HGC27 cells respectively and detected the cell cycle distribution by flow
cytometry (Figure 5A and B). The results showed that the proportion of cells in G2 phase was
significantly increased in the puerarin-treated group compared with the DMSO control group, indicating that puerarin
was able to induce cells to arrest in G2 phase. To further verify this phenomenon, we examined the expression levels
of cell cycle-related proteins (CDK1, CDK4, CyclinA and CyclinB) in the DMSO group, the 1 mM group and the 2 mM group
by using the Western blot technique (Figure 5C and D). The results showed that puerarin treatment
significantly down-regulated the expression of CDK1 and CyclinB, while affecting the expression pattern of CDK4 and
CyclinA. In addition, the effect of puerarin on cell cycle regulation was further confirmed by detecting the content
of CDK4 in HGC27 cells by cellular immunofluorescence assay (Figure 5E). In summary, the above
experimental results collectively indicated that puerarin promotes cell arrest in G2 phase by regulating the
expression of cell cycle-related proteins, thus inhibiting the transformation of cells to M phase.
Figure 5. Effects of Puerarin on the cell cycle of gastric cancer (A)The cell cycle distribution of AGS gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was examined by flow cytometry to analyse the changes in the proportions of cells in the G1, S and G2 phases. (B) The cell cycle distribution of HGC27 gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was examined by flow cytometry to analyse the changes in the proportions of cells in the G1, S and G2 phases. and G2 phases. (C) The expression levels of AGS cell cycle-related proteins (CDK1, CDK4, CyclinA and CyclinB) and the endogenous protein GAPDH were detected in DMSO (control group), 1 mM and 2 mM Pueraria Mirifica-treated groups using Western blot. (D) The expression level of expression levels of HGC27 cell cycle-related proteins (CDK1, CDK4, CyclinA and CyclinB) and the endogenous reference protein GAPDH in the DMSO (control), 1 mM and 2 mM geraniol-treated groups. (E) Cellular immunofluorescence was used to detect the expression of CDK4 in HGC27 gastric cancer cells.
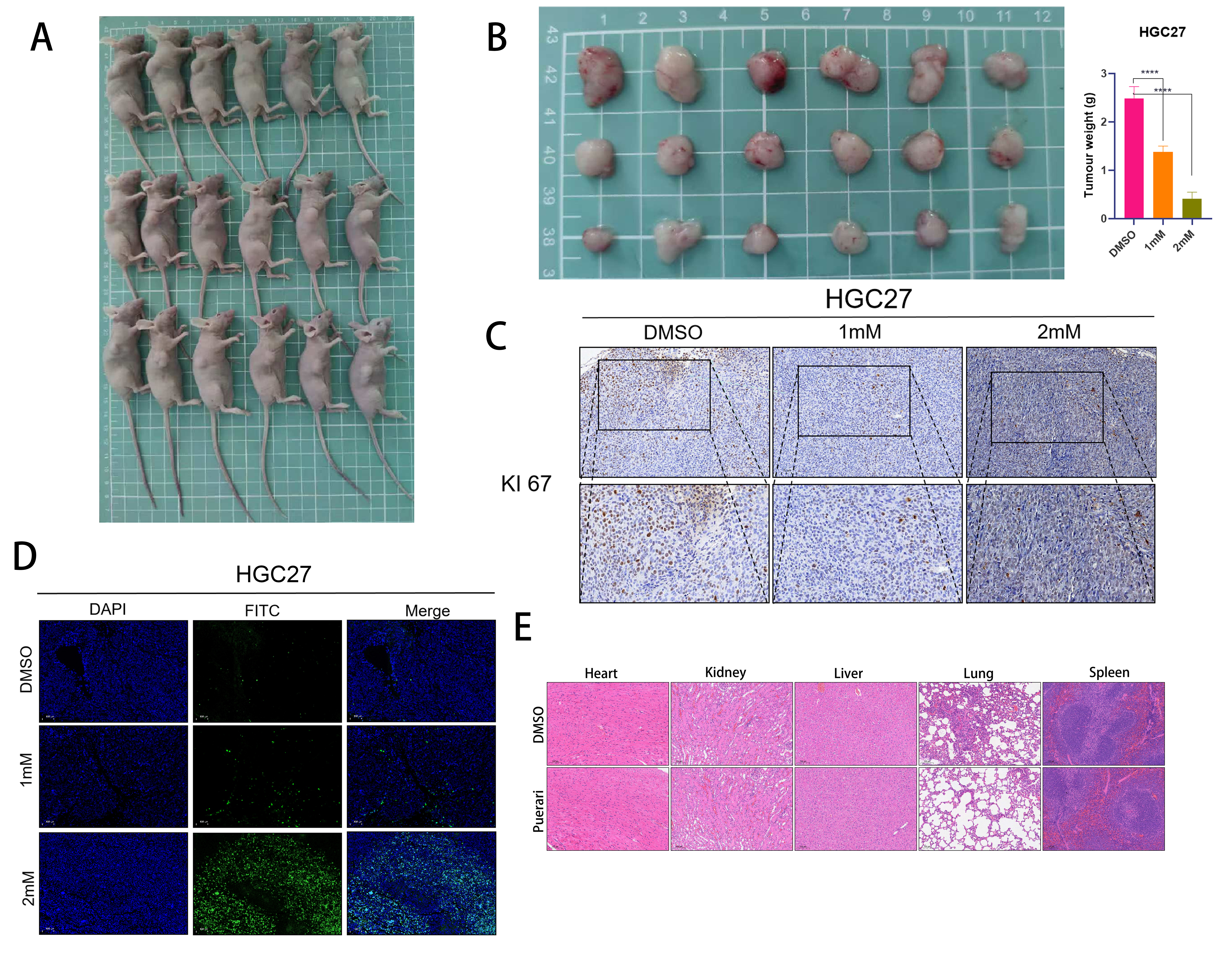
Effect of puerarin on tumours in vivo
To investigate the effect of puerarin on tumour growth in vivo, we
performed subcutaneous transplantation tumour experiments. First, 1 × 10⁷ HGC27 cells were injected into the axillary
region of mice, and one week later, the mice were randomly divided into three groups: the DMSO control group (injected
with saline), the 1 mM group (injected with 20 μg of puerarin), and the 2 mM group (injected with 40 μg of puerarin).
The experimental results showed that puerarin treatment significantly inhibited the growth of tumours in vivo,
indicating its significant anti-tumour activity (Figure 6A and B). To further assess the effect of
puerarin on the proliferation and apoptosis of tumour cells, we sectioned the tumour tissues and detected the
proliferation and apoptosis levels of tumour cells using KI-67 staining and TUNEL staining respectively (Figure 6C). The results showed that the proportion of KI-67-positive cells was significantly reduced in the puerarin-treated
group, whereas the proportion of TUNEL-positive cells was significantly increased, indicating that puerarin was able
to inhibit the proliferation of tumour cells and induce their apoptosis (Figure 6D). In addition, to
assess the potential toxicity of puerarin on the major organs of mice, we performed HE staining analysis on the heart,
liver, kidney, lung and spleen of mice in the normal and treated groups (Figure 6E). The results
showed that no significant pathological changes were observed in the major organs of mice in all groups, indicating
that puerarin had no significant organ toxicity at the experimental dose. In conclusion, this study confirmed that
puerarin could effectively inhibit the growth of tumours in vivo without showing obvious organ toxicity, which
provides an important experimental basis for its further development as an anti-tumour drug.
Figure 6. Effects of Puerarin in rats (A)At the end of the experiment, nude mice in the DMSO control group, 1 mM group and 2 mM group were executed to collect tumour tissues and major organs for subsequent analysis. (B) The inhibitory effect of puerarin on tumour growth was assessed by measuring the volume of tumours in nude mice in the DMSO control group, 1 mM group and 2 mM group. (C) Sections of tumour tissues in the DMSO control, 1 mM and 2 mM groups were subjected to KI67 staining for the detection of tumour growth. KI67 staining was performed to detect the proliferation level of tumour cells to clarify the effect of puerarin on tumour cell proliferation. (D) TUNEL staining was performed on tumour tissue sections from DMSO control, 1 mM and 2 mM groups to detect the apoptosis level of tumour cells, to further validate the apoptosis-inducing effect of puerarin on tumour cells. (E) Hepatitis B and Hepatitis C staining was performed on hearts, livers, kidneys, lungs, and spleens of nude mice in DMSO control and 2 mM groups to assess the inhibitory effect of puerarin on tumour growth. , lungs and spleens were subjected to HE staining to assess the potential toxicity of gerberellin to major organs and ensure its safety.
Discussion
Puerarin is a natural isoflavonoid, one of the main active components in Pueraria lobata. It has a variety of pharmacological effects, including antioxidant, anti-inflammatory, cardiovascular protection, neuroprotection, etc.[19] In recent years, it has been found that puerarin is closely related to cancer, and that it affects the proliferation, apoptosis, invasion, and metastasis of tumour cells through a variety of mechanisms, possesses both antioxidant and anti-inflammatory effects and may indirectly inhibit tumourigenesis and progression. For example, in breast cancer puerarin inhibits the proliferation and invasion of breast cancer cells by inhibiting the PI3K/AKT and MAPK signalling pathways[19,21]. In hepatocellular carcinoma, Puerarin inhibits the proliferation and metastasis of hepatocellular carcinoma cells by modulating the Wnt/β-catenin and PI3K/AKT signalling pathways. In animal models, puerarin significantly inhibited the growth of hepatocellular carcinoma. Meanwhile, Puerarin is also closely related to cancer treatment. Experimental studies have shown that Puerarin can enhance the sensitivity of tumour cells to chemotherapeutic drugs (e.g., cisplatin, 5-fluorouracil), and reduce the dosage and side effects of chemotherapeutic drugs. For example, in lung and gastric cancer studies, puerarin showed synergistic anti-tumour effects in combination with chemotherapeutic drugs. Through its antioxidant and anti-inflammatory effects, puerarin reduces oxidative stress and inflammation in tumour patients, thereby improving their quality of life. Recent studies highlight that dysregulation of the cell cycle plays a central role in gastric cancer, closely linked to multi omics molecular subtypes. A G2/M phase related PDEGs signature can predict survival and immunotherapy response, with key genes such as F2R associated with oxaliplatin resistance, while natural products and epigenetic modulation (e.g., p21 upregulation) induce G2/M arrest, while axes involving UBE2C and Hip2 regulate proliferation and metastasis, offering novel targets for precision therapy[26]. The aim of this experiment was to investigate the effect of puerarin on gastric cancer and its potential mechanism. Firstly, we selected two common gastric cancer cell lines (HGC27 and AGS) and determined the optimal administration concentration of puerarin within 24 hours by a CCK-8 assay, and found that puerarin had a significant inhibitory effect on the proliferation of gastric cancer cells. To further validate this result, we used EdU staining and plate cloning experiments, both of which confirmed that puerarin could effectively inhibit the proliferation ability of gastric cancer cells. Given the high metastatic characteristics of gastric cancer, we explored the effect of puerarin on the invasive ability of gastric cancer cells by transwell assay. The results showed that puerarin significantly inhibited cell invasion. In order to further investigate the mechanism of puerarin, we performed transcriptome sequencing on the blank and puerarin-treated groups. Transcriptome sequencing analysis revealed that puerarin inhibited cell proliferation primarily by affecting the expression of cell cycle-related genes. Therefore, we further detected the changes in cell cycle by flow cytometry and found that, compared with the control group, the additive group showed a significantly increased proportion of cells in G2 phase and a significantly decreased proportion in G1 phase. This suggests that puerarin may inhibit tumour cell proliferation by blocking the cell cycle. In addition, the transcriptome sequencing results also showed that puerarin was able to induce apoptosis. To verify this, we detected key proteins of apoptosis (e.g., Bax, Bcl-2 and Caspase-3) by flow cytometry and Western blotting, and the results further confirmed that puerarin could promote apoptosis of gastric cancer cells. Taking the above experimental results together, we systematically elucidated the inhibitory effect of puerarin on gastric cancer cells and its potential mechanism in terms of cell proliferation, invasion, cycle arrest, and apoptosis. Although we performed multiple experiments to investigate the cell cycle and apoptosis, further investigation is still needed as to the way in which puerarin leads to cell cycle changes. In order to further investigate the anti-tumour effects of puerarin in vivo, we carried out animal experiments in which the experimental animals were divided into the treatment group and control groups, and the effects of puerarin on tumour growth were investigated by constructing a subcutaneous tumour-forming model in nude mice. The experimental results showed that the tumour volume of the drug-added group was significantly smaller than that of the control group, suggesting that puerarin has an obvious tumour-suppressing effect in vivo. To further verify this result, we sectioned the tumour tissues of nude mice and analysed the proliferative activity of tumour cells by KI-67 immunohistochemical staining, and found that the proportion of KI67-positive cells in the drug-added group was significantly reduced, which further confirmed the inhibitory effect of puerarin on tumour proliferation. Although we preliminarily verified the in vivo anti-tumour effect of puerarin through the nude mice tumour-forming assay, this study has not yet been subjected to clinical trials, and therefore the actual efficacy and safety of puerarin in humans cannot be assessed. In addition, future studies could further explore the combined effects of puerarin and chemotherapeutic agents (e.g., cisplatin) to assess whether it can enhance the effects of chemotherapy or reduce the side effects of chemotherapeutic agents, thus providing new strategies for the combination therapy of gastric cancer. In summary, the present study demonstrated that puerarin inhibited the proliferation of tumour cells by regulating the cell cycle, providing an experimental basis for its use as a potential anticancer drug. However, further clinical studies are needed to assess the efficacy and safety of puerarin in humans, as well as to explore its synergistic effects with other chemotherapeutic agents. In addition, more in-depth studies on the mechanism of action of puerarin in gastric cancer are also needed to reveal the molecular mechanisms behind it and to provide a new theoretical basis and a direction for drug development in gastric cancer treatment.
Conclusion
Our findings demonstrate that puerarin significantly suppresses malignant biological behaviors in gastric cancer cells. Transcriptome sequencing analysis revealed that puerarin inhibits cell proliferation by blocking cell cycle progression. Furthermore, in vivo experiments confirmed that puerarin effectively reduces the tumorigenic capacity of gastric cancer cells in a nude mouse subcutaneous xenograft model.
Abbreviations
EdU: 5-Ethynyl-2'-deoxyuridine; WB:Western Blot; DMSO: dimethylsulfoxide; GAPDH: glyceraldehyde-3-phosphate; CCK8: Cell Counting Kit-8; IC50: Half maximal inhibitory con-centration; Rpm: Revolutions per minute; DAPI: 4', 6-diamidi-no-2-phenylindole; 594: Fluorescein 594.
Declarations
Author Contributions
Jianye Han: Experimental design and execution, Writing original draft. Weiwei Yuan: Data management. All authors read and approved the final manuscript.
Acknowledgments
Not Applicable.
Funding Information
Not Applicable.
Ethics Approval and Consent to Participate
This study was approved by the Animal Ethics Committee of Anhui Medical University (Protocol No. 2024123). All procedures were conducted in strict compliance with the Guidelines for Ethical Review of Laboratory Animal Welfare in China (GB/ T 35892-2018) and the ARRIVE 2.0 guidelines to ensure the ethical treatment of animals. No human tissues were used.
Competing Interests
The authors declare that they have no existing or potential commercial or financial relationships that could create a conflict of interest at the time of conducting this study.
Data Availability
All data needed to evaluate the conclusions in the paper are present in the paper or the Supplementary Materials. Additional data related to this paper may be requested from the authors.
References
 Figures
Figures References
References Peer
Peer Information
InformationFigure 1. Spatial structure of Puerarin and toxic effects of two gastric cancer cells (A)Two-dimensional chemical structure of puerarin. (B) Three-dimensional molecular structure of puerarin. (C) The effect of different concentration gradients of puerarin treatment on the viability of HGC27 cells over 24 h was detected by CCK-8 method. (D) The effect of different concentration gradients of puerarin treatment on the viability of AGS cells over 24 h was detected by CCK-8 method. (E) The concentrations of DMSO (control), 1 mM, and 2 mM were used to treat HGC27 gastric cancer cells, respectively. of Puerarin were treated with HGC27 gastric cancer cells and cell viability was determined by CCK-8 assay at 0, 12, 24, 36 and 48 h time points. (F) AGS gastric cancer cells were treated with DMSO (control), 1 mM and 2 mM concentrations of Puerarin, respectively, and cell viability was determined by CCK-8 assay at 0, 12, 24, 36 and 48 h time points.
Figure 2. Puerarin inhibited the proliferation and invasion of gastric cancer (A)The fluorescence positivity rate of AGS cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was detected by EdU assay to evaluate the effect of the drug on the proliferative viability of the cells. (B) The fluorescence positivity rate of HGC27 cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was detected by EdU assay to further evaluate the effect of Puerarin on the proliferative ability of the cells. (C) The monoclonal formation ability of AGS and HGC27 cells under different concentrations of puerarin treatment was analysed by colony formation assay to clarify the dose-dependent effect of the drug on the cell proliferation function. (D) The effects of different concentrations of puerarin treatment on the invasion ability of AGS and HGC27 cells were investigated by using Transwell assay to reveal the potential mechanism of the drug's action in inhibiting cell migration and invasion. (E) The effects of different concentrations of puerarin treatment on the invasion ability of AGS and HGC27 cells were investigated by using Transwell assay. potential mechanism of action in inhibiting cell migration and invasion.
Figure 3. Analysis of the transcriptome sequencing results revealed the significant effects of Puerarin on the gene expression profiles of gastric cancer cells. (A) RNA sequencing results showed that more than 2,000 significantly up-regulated genes and more than 1,000 significantly down-regulated genes existed in the Puerarin-treated group compared with the control group. (B) KEGG dot plot enrichment analysis further demonstrated that the differentially expressed genes were significantly enriched in the DNA replication and cell-cycle related pathways. (C) KEGG histogram enrichment analysis results were consistent with the dot plot, which further verified that DNA replication and cell cycle pathways in the mechanism of action of puerarin. (D) GO functional annotation histogram showed that the differentially expressed genes were significantly enriched in cell cycle-related functional categories, which further supported the regulatory effect of puerarin on cell cycle progression. (E) In addition, the sequencing results also showed that puerarin significantly affected the apoptosis-related pathway, suggesting that it may inhibit the growth of gastric cancer cells by inducing cell apoptosis. growth of gastric cancer cells by inducing apoptosis.
Figure 4. Detection of apoptosis by flow and WB experiments (A)Detection of apoptosis of AGS gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups by flow cytometry. (B) Detection of apoptosis of HGC27 gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups by flow cytometry. (C) Detection of apoptosis in DMSO (control group), 1 mM and 2 mM expression levels of AGS apoptosis-related proteins (BAX, BCL2, Caspase3) and the internal reference protein GAPDH in the Puerarin-treated groups. (D) The expression levels of apoptosis-related proteins (BAX, BCL2, Caspase3) and the internal reference protein GAPDH in HGC27 cells in the DMSO (control group), the 1 mM and the 2 mM Puerarin-treated groups were examined by Western blotting technique. protein GAPDH in the DMSO (control) and 2 mM Puerarin-treated groups
Figure 5. Effects of Puerarin on the cell cycle of gastric cancer (A)The cell cycle distribution of AGS gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was examined by flow cytometry to analyse the changes in the proportions of cells in the G1, S and G2 phases. (B) The cell cycle distribution of HGC27 gastric cancer cells in DMSO (control group), 1 mM and 2 mM Puerarin-treated groups was examined by flow cytometry to analyse the changes in the proportions of cells in the G1, S and G2 phases. and G2 phases. (C) The expression levels of AGS cell cycle-related proteins (CDK1, CDK4, CyclinA and CyclinB) and the endogenous protein GAPDH were detected in DMSO (control group), 1 mM and 2 mM Pueraria Mirifica-treated groups using Western blot. (D) The expression level of expression levels of HGC27 cell cycle-related proteins (CDK1, CDK4, CyclinA and CyclinB) and the endogenous reference protein GAPDH in the DMSO (control), 1 mM and 2 mM geraniol-treated groups. (E) Cellular immunofluorescence was used to detect the expression of CDK4 in HGC27 gastric cancer cells.
Figure 6. Effects of Puerarin in rats (A)At the end of the experiment, nude mice in the DMSO control group, 1 mM group and 2 mM group were executed to collect tumour tissues and major organs for subsequent analysis. (B) The inhibitory effect of puerarin on tumour growth was assessed by measuring the volume of tumours in nude mice in the DMSO control group, 1 mM group and 2 mM group. (C) Sections of tumour tissues in the DMSO control, 1 mM and 2 mM groups were subjected to KI67 staining for the detection of tumour growth. KI67 staining was performed to detect the proliferation level of tumour cells to clarify the effect of puerarin on tumour cell proliferation. (D) TUNEL staining was performed on tumour tissue sections from DMSO control, 1 mM and 2 mM groups to detect the apoptosis level of tumour cells, to further validate the apoptosis-inducing effect of puerarin on tumour cells. (E) Hepatitis B and Hepatitis C staining was performed on hearts, livers, kidneys, lungs, and spleens of nude mice in DMSO control and 2 mM groups to assess the inhibitory effect of puerarin on tumour growth. , lungs and spleens were subjected to HE staining to assess the potential toxicity of gerberellin to major organs and ensure its safety.
Peer-review Terminology
Identity transparency: Single anonymized
Reviewer interacts with: Editor
Details
This is an open access article under the terms of the Creative Commons Attribution License(http://creativecommons.org/licenses/by/4.0/), which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Publication History
Received 2025-10-11
Accepted 2026-12-23
Published 2026-02-10